接合生殖
接合生殖
大腸桿菌有兩種性別系,當不同性系的個體雜交時,陽性菌體細胞接觸陰性菌體細胞,兩個細胞側面形成接合管,陽性系細胞的DNA通過接合管流入陰性系細胞內,形成接合子;接合子經過細胞分裂形成兩個大腸桿菌。水綿接合生殖時,陽性接合細胞內全部原生質通過接合管到達陰性接合細胞,由兩個細胞的原生質融合而生成接合子;接合子經過減數分裂生成有性孢子各2個,有性孢子萌發為水綿的營養體。草履蟲接合生殖時,每個蟲體大核消失,小核減數分裂生成4個核,其中3個核消失,留下的一個核分成動核和靜核;動核通過接合膜交換,分別與對方的靜核融合;接合結束后,兩個蟲體分開,各自經歷三次核分裂和兩次胞質分裂,形成4個新個體.
草履蟲的接合生殖
草履蟲種內存在不同的繁殖群(syngen),每一繁殖群內存在不同的接合型(matingtype),只有同繁殖群內不同的接合型之間才能發生接合,進行接合生殖。在不同繁殖群之間,不同的接合型間一般不發生接合。有的草履蟲根據繁殖群之間的這種有性生殖的隔離現象,這些繁殖群已被上升為不同的姊妹種,如原來的雙小核草履蟲(parameciumaurelia)的1個種已分成14個姊妹種(siblingspecies)。其它草履蟲仍保留繁殖群(syngen)這一種下的分類單位。根據草履蟲種類的不同,每一姊妹種或繁殖群內接合型的組成也不同,如雙小核算履蟲((P.aurelia)中的每一姊妹種和尾草履蟲(P.caudatum)及多小核草履蟲(P.multmicronucleatum)中的每一繁殖群內只有兩個可發生接合的接合型,而綠草履蟲(P.bursaria)和旋毛草履蟲(P.trichium)的每一繁殖群內則有多個接合型,其中任何兩型之間都可接合。因此接合型不同於其它動物的“性”。因為“性”不可能有多個,又因草履蟲接合時每一個體內都產生兩種原核,相當於精核和卵核,接合時精核互換與對方的卵核受精,故草履蟲屬於雌雄同體的動物,個體之間無性別之分。草履蟲的不同接合型屬於不同無性系(clone),無性系追溯到它的起源也可稱之為大核系(caryonide),因為草履蟲的無性繁殖主要是靠大核的不間斷的分裂。接合生殖時由小核產生受精作用重建新的大核,不同草履蟲接合一次建成的新大核數目不同,一般每一接合后體不是建成4個就是建成2個,接合后體通過整個細胞的分裂各分得單一的新大核,從此由新大核通過無性分裂傳下的後代群就叫大核系,亦即無性系。草履蟲的不同接合型就分佈在不同的大核系也即無性系中。
(二)獲得草履蟲接合型的方法
獲得草履蟲純的不同接合型有多種方法,這裡只介紹最容易的方法,即無性系法。任何草履蟲在它的繁殖地都至少存在一個繁殖群內的兩個或多個接合型,它們由不同的無性系來體現。將由自然環境中採得的草履蟲單個分離,用原地過濾並煮沸放冷的池水洗去一切其它生物,將這些草履蟲放入凹玻片的凹坑內,加入稻草湯培養液(製法見后),放入 20℃—25℃下培養,數日後即繁殖成群,再移入試管培養。蟲體增多后要每日至少加一次新培養液,以保持其旺盛的無性繁殖。這樣建起的多個無性系,必然包含相對的接合型。在適宜條件下,將這些無性系中的每兩個取樣相混,凡能發生接合的兩個無性系就是兩個相對接合型。找出相對接合型后,永遠保持其食物充分和生長旺盛,就隨時可取樣相混,立即獲得接合對。如欲保存以後備用,可放入4℃—6℃冰箱中,每半月到1月加一次新培養液,可長期保存接合型。用時升溫加食,經過幾天快速繁殖后,相混仍可產生接合反應。
(三)控制草履蟲接合的方法
草履蟲能否進行接合生殖取決於四個因素,即接合型,成熟度、食物和溫度。
如前所述,革履蟲種內或繁殖群內存在不同的接合型,只有相對接合型之間才能有接合反應,進行有性生殖。屬同一接合型的不同無性系之間一般不能接合。
草履蟲的一個大校系的一生可分為四個階段,即未成熟期、成熟期、衰老期和死亡期。未成熟期是指新生的大核系只經較少次數的無性分裂階段。處於此期的草履蟲即使是屬於相對的兩個接合型,相混后也不會有接合反應,即使有百分率也非常低。只有達到成熟以後,相對接合型之間才可能出現接合反應,而且有較高的接合率。處於衰老期的蟲體由於代謝及生理上的原因,雖屬相對接合型的個體相遇,接合率也會明顯降低,而且接合后體的成活率也顯著下降。
達到成熟期的相對接合型能否相混后立即接合還取決於蟲體的營養狀況。過度飽食或過度飢餓都不能出現接合反應。因此,想得到理想的接合材料必須控制食物。只有旺盛繁殖后,由飽食轉入飢餓的數小時內,兩型相混才會立即發生接合。發生接合的溫度在15℃—30℃之間,20℃—25℃為最佳。上述四個因素是控制接合不可忽視的。
草履蟲培養液可按下法制出:取10克稻草或麥桿加水1000毫升,煮沸過濾,放涼後接種產氣桿菌屬(Aerobacter)的細菌,放入30℃下培養3天,稻草湯變混濁后即可使用(如無產氣桿菌,也可在空氣下暴露3天後使用)。將已建起的無性系用此培養液培養數日,高速繁殖后都可達到成熟期,當加食后待培養物開始變為清徹透明時,即將達到飽食轉入飢餓的時期,這時將屬於相對接合型的兩個培養物相混,立即或在數小時內發生接合。接合初期的反應是幾個甚至幾十個蟲體相互粘著、聚團。過1小時以後將出現大量的接合對,開始接合生殖的內部過程。用這些方法既可找出相對的接合型,也可難持接合型,隨時相混即能獲得供實驗用的接合材料。
(四)觀察草履蟲接合過程的方法
由上述方法獲得理想的接合材料,用定期壓片(根據情況每隔10—30分鐘),石炭酸品紅染色的方法做臨時裝片,這樣就可以觀察到接合過程中整個核的變化情況。具體方法如下:1.用離心機的最低轉數將接合材料濃縮,吸1滴置於載玻片的中央或者用微吸管吸幾個接合對於載玻片中央(切記水不要過多),在蟲體周圍點4點漿糊,然後加上蓋片,用鑷子或鉛筆尖在蓋片的四角向下壓直至蟲體只能緩慢移動為止。2.在蓋片的一側滴加石炭酸品紅染液,在對側用吸水紙吸去多餘的染液,染15分鐘后即可觀察。這樣的臨時裝片也可經梯度酒精脫水,二甲苯透明,樹膠封片,做成永久裝片。蓋片在脫水過程中脫落,可將載片和蓋片分別封成兩張片子,上面都會留有接合的蟲體。
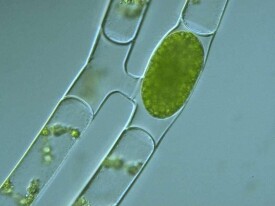
水綿的有性生殖為接合生殖。其中又包括梯形接合、側面和直接側面接合3種類型,但以梯形接合為最常見。接合生殖多於春季和秋季發生,這時絲狀體的顏色也由綠變為黃綠,
其接合生殖過程是:首先為2條水綿絲狀體并行靠近,在兩條絲狀體的細胞相對的一側各產生1個突起。繼而兩相對突起伸長,頂端接觸,端壁融解形成1條連通的管子,叫做接合管。同時,各個細胞內的原生質體濃縮,形成1個配子。其中1條絲狀體的每個細胞中的配子,以變形蟲式運動通過接合管移入另1條相對細胞中,與其中的1個配子融合,產生1個卵形或橢圓形的合子,也稱接合孢子。這樣,1條絲狀體的細胞都變空,另1條絲狀體的細胞中都各有1個合子。空的絲狀體可視為雄性,有合子的絲狀體可視為雌性。以上生殖過程就叫做接合生殖。由於在兩條絲狀體之間有很多橫向的接合管,外觀上看頗像個梯子,故稱做梯形接合。以後合子分泌產生厚壁,藻體死亡崩解后,合子沉入水底休眠。條件適宜時萌發,合子核首先經過減數分裂,所產生的3個單倍體核退化,僅1個發育,最後產生1條新的水綿絲狀體