蛋白質合成體系
蛋白質合成體系
原核生物一個mRNA帶有功能相關的幾種“蛋白質”的編碼信息,稱多順反子(幾個基因的複本);真核生物一個mRNA一般只帶一種蛋白質的編碼信息,稱單順反子。mRNA的生成要經加工,尤其是真核生物細胞,這就造成mRNA的序列和DNA序列間沒有完整的一對一的關係。遺傳密碼(genetic code)是規定mRNA的核苷酸序列翻譯成多肽鏈氨基酸序列的一套法則,也就是mRNA的核苷酸序列和多肽鏈氨基酸序列的共線性關係。
1961年Crick及其同事通過研究噬菌體基因的移碼突變推測三聯體密碼子是非重疊、無標點的。Nirenberg等用人工合成的mRNA在無細胞蛋白質合成系統中尋找氨基酸與三聯體密碼子的對應關係。Khorana和他的同事用化學合成結合酶促反應,合成含有2、3、4核苷酸重複序列的多聚核苷酸,以此為模板找出各氨基酸的密碼子。技術上的突破來自人工合成的三核苷酸能與對應的氨醯-tRNA一起結合在核糖體上,由此確定絕大多數密碼子。1966年全部64個密碼子破譯,其中AUG編碼甲硫氨酸,又是起始密碼;UAA、UAG、UGA3個是終止密碼,不編碼氨基酸;還有 61個編碼一特定的氨基酸。
遺傳密碼特點:①連續性,指密碼子必須按5′→3′方向三個一組讀碼框往下閱讀,無標點、不重疊、不跳格。正確的讀碼框的確立是由核糖體識別在編碼序列開頭處的起始密碼AUG;②簡併性,是指同一種氨基酸有兩個或更多密碼子的現象。編碼同一氨基酸的密碼子稱為同義密碼子,通常只在第3位鹼基上不同,這樣可減少有害突變。密碼子第3位鹼基與tRNA反密碼子不嚴格遵從鹼基配對規律(擺動鹼基配對),如tRNA反密碼子第一位的I(由A轉變而來)可與mRNA密碼子第3位鹼基U、C、A形成配對,U可對應A、G,因而密碼子第3個位置又稱擺動位置;③通用性,即所有生物基本共用同一套遺傳密碼。線粒體以及少數生物基因組的密碼子有變異(如在酵母、哺乳動物、果蠅中,AUA=Met而非Ile,UGA=Trp而非終止碼。)
蛋白質合成體系
氨醯-tRNA合成酶具有絕對專一性,對L-氨基酸、tRNA兩種底物能高度特異識別。大腸桿菌丙氨酸tRNA的氨基酸接受臂上的G3·U70鹼基對決定負載Ala的專一性。精氨酸-tRNA(A20),異亮氨酸-tRNA(G5·G69),酵母苯丙氨酸-tRNA(G20,G34,A35,A36)。由於氨基酸和tRNA正確結合,而tRNA又和mRNA、核糖體準確配對,這就確保遺傳信息傳遞的穩定。氨醯-tRNA合成酶與tRNA之間的相互作用和tRNA分子中某些鹼基或鹼基對決定著攜帶專一氨基酸的作用組成tRNA分子第二套密碼系統。
1950年P.Zamecnik將放射性同位素標記的氨基酸注射到小鼠體內,經短時間后,取出肝臟,製成勻漿,離心,分成核、線粒體、微粒體及上清液組分,發現微粒體中的放射性強度最高,再處理微粒體,將核糖體從內質網中分離出,發現核糖體的放射強度比微粒體高7倍。
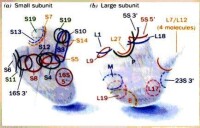
有70S和80S兩種,均由大小不同的兩個亞基組成。70S核糖體存在於原核細胞和真核細胞的線粒體和葉綠體中,其30S小亞基含有一個16SrRNA和21種不同的蛋白質(稱S蛋白),50S大亞基含有一個23SrRNA、5SrRNA和34種蛋白質(L蛋白)。80S核糖體存在於真核細胞,其40S小亞基含有一個18SrRNA和34種S蛋白,60S大亞基含有28SrRNA、5SrRNA、5.8SrRNA各一分子和49種L蛋白。在通常情況下,核糖體的大小亞基遊離於細胞質基質中,只有當小亞基與mRNA結合后,大亞基才與小亞基結合形成完整的核糖體。
核糖體上有兩個tRNA結合的位點:A位點是氨醯tRNA結合位,P位點是肽醯tRNA結合位。50S亞基上有一個GTP水解位點,為氨醯-tRNA移位提供能量;兩亞基接觸面空隙有結合mRNA的位點,還有與起始因子、延伸因子、釋放因子及各種酶相結合的位點,mRNA和合成的新生多肽鏈通過外出孔進入膜腔。

基本信息
- 中文名
- 蛋白質合成體系
- 外文名
- genetic code
- 內容
- mRNA核苷酸和多肽鏈氨基酸
- 稱
- 多順反子