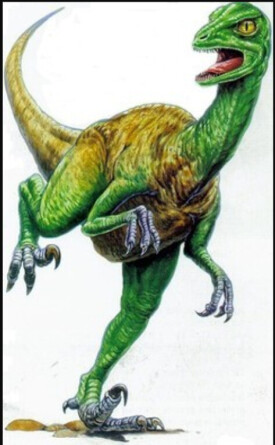
傷齒龍
蜥臀目傷齒龍科動物
傷齒龍生存於晚白堊紀,約7500萬年前到6500萬年前,最初是因為它尖銳的牙齒而得名。剛開始人們首先認為它是一種蜥蜴,然後又把它當作一種長相呆笨的恐龍,後來把它的骨骼組合起來之後,才發現以前的認識和理解幾乎全是錯誤的。就身體和大腦的比例來看,傷齒龍的大腦是恐龍中最大的,而且它的感覺器官非常發達,因而被認為是最聰明的恐龍。有些科學家甚至認為它可能和鴕鳥智商相近,那將比現生的任何爬行動物都要聰明。袋鼠的IQ大約在0.7。而傷齒龍的IQ高達5.3!好傢夥,真是高智力啊。傷齒龍可能和今天鳥類的智力有些相似。今天的鳥類極為聰明:最聰明的鳥能訓練開一些玩笑甚至模仿人類的語言。加拿大古動物學家戴爾羅素就設想,如果6500萬年前沒有那場大災難,迅猛龍的近親奔龍,就完全可能進化成為代替人類的一種動物——“恐龍人”,成為地球的主宰。這就是曾經風行一時的“恐人學說”。
傷齒龍起初被認為是種掠食動物,主要以較小型動物為食,復原圖例如無脊椎動物或哺乳類。這個論點是根據傷齒龍的手盜龍類特徵(例如:腳部的鐮刀狀趾爪、類似鳥類的可抓握手臂),以及好的深度知覺。然而,有些證據顯示傷齒龍可能是雜食性或草食性動物。頜部有著寬廣、U形的聯合線,類似鬣蜥;鬣蜥是種已適應草食性的蜥蜴。另外,傷齒龍的牙齒呈葉狀,有著大型鋸齒狀邊緣,如同草食性恐龍,而且牙齒短而寬廣,側邊有磨損面。在1998年,湯瑪斯·霍爾茲(Thomas R. Holtz)認為傷齒龍的可抓握手臂、較大型腦部、深度知覺,跟雜食性/草食性的靈長類、雜食性的浣熊具有共同的特性。
一個2008年的研究,分析了來自北美洲各地的傷齒龍牙齒化石,發現阿拉斯加州北坡地區的牙齒,大於較南地區的牙齒。這顯示北坡地區的傷齒龍普遍成長至較大的體型。研究人員推測,這是因為沒有暴龍科生存於北坡地區,所以該地區的傷齒龍面臨較少的競爭者,可以成長至較大的體型。該研究也分析牙齒的磨損模式,認為傷齒龍是以質地較軟的食物為食,而無法咬裂骨頭或無脊椎動物的外骨骼、以及堅硬的植物。研究人員推測,傷齒龍主要以肉類為食。
根據化石的年齡鑒定,傷齒龍死亡時的平均年齡為3到5歲。
恐龍可能會進化成“恐龍人在很長時間以來,科學家都一直認為恐龍是種笨拙、冷血,隨時等待滅絕的動物。然而隨著更多科學發現最近浮出水面,譬如考古學家在地球兩極地區都發現了恐龍化石,從而迫使科學家徹底改變了以前對恐龍的“偏見”。美國明尼蘇達州科學博物館古生物學家克里斯蒂·科里·羅傑斯稱,一些恐龍已經具有足夠的能力,可以適應地球演變過程中發生的氣候劇變,這意味著如果恐龍沒有遭遇6500萬年前的那場滅頂之災,它們將可以繼續倖存下來,甚至度過冰河時代。加拿大阿爾伯達大學古生物學家菲爾·科里說稱,如果6500萬年前的那顆小行星並沒有撞中地球,那麼他相信恐龍至今仍將是地球的主宰。科學家認為,假如人類能夠和恐龍生活在同一個時代,人類沒有牛羊可以放牧,也許可以養殖一些恐龍。而1.2米長、2.5公斤重的溫順的奇齒龍,則可以成為人類的完美寵物。科學家認為,在恐龍瀕臨滅絕的時代,當時最高級、最聰明的恐龍大概要數傷齒龍,據美國俄亥俄州大學古生物學家拉里·惠特默稱,傷齒龍“就像狐狸一樣狡猾”,它們個子很小,直立行走,喜歡群居。通過研究它們的大腦容量,惠特默發現它們不但擁有良好的視力,甚至還擁有潛在的解決問題的能力。有科學家相信,傷齒龍很可能會沿著靈長類或人類的發展方向進化,“最後進化出智慧的恐龍人”。
白堊紀(CretaceousPeriod,Cretaceous)中生代最後的一個紀。白堊紀(Cretaceusperiod)是中生代的最後一個紀,始於距今1.37億年,結束於距今6500萬年,其間經歷了7000萬年。無論是無機界還是有機界在白堊紀都經歷了重要變革。位於侏羅紀之上、新生界之下。白堊紀是中生代地球表面受淹沒程度最大的時期,在此期間北半球廣泛沉積了白堊層,1822年比利時學者J.B.J.奧馬利達魯瓦將其命名為白堊系。白堊層是一種極細而純的粉狀灰岩,是生物成因的海洋沉積,主要由一種叫做顆石藻的鈣質超微化石和浮遊有孔蟲化石構成。在這一時期,大陸之間被海洋分開,地球變得溫暖、乾旱。開花植物出現了,與此同時,許多新的恐龍種類也開始出現,包括像食肉牛龍這樣的大型肉食性恐龍,像戟龍這樣的甲龍類成員以及像賴氏龍這樣的植食性鴨嘴龍類。恐龍仍然統治著陸地,像飛機一樣的翼龍類,例如披羽蛇翼龍在天空中滑翔,巨大的海生爬行動物,例如海王龍統治著淺海。但最早的蛇類、蛾、和蜜蜂以及許多新的小型哺乳動物也在這一時期出現了。白堊紀時,南美洲與非洲大陸之間的裂谷迅速張開形成南大西洋,到末期已加寬到約3000千米。北大西洋裂谷位於格陵蘭和北美東側,隨著北美洲向西漂移,裂谷在擴大。特提斯海把歐亞大陸與非洲分開,中南歐和中近東的許多國家當時都處於海侵中。白堊紀時氣候比較溫暖,未見極地冰蓋跡象。當時大部分地區雨量充沛,氣候濕潤,近海及濱海地帶形成豐富的石油、煤、天然氣和油頁岩礦床,如美國得克薩斯州、墨西哥、波斯灣、北非和俄羅斯的許多大油田,又如中國松遼平原上白堊統的大慶油田,東北和內蒙古下白堊統的許多大煤田。在一些氣候乾旱炎熱的地區,如中國南方晚白堊世的西南湖群和雲夢澤水系,有巨厚的膏鹽礦床沉積。此外,在雲南白堊紀紅層中,含銅砂礦床品位高,規模大,易於開採和冶鍊,是中國有名的銅鄉。
傷齒龍是由約瑟夫·萊迪(Joseph Leidy)在1856年所命名,最初的拼法為Troödon。在1876年,由H. E. Sauvage正式修正為目前的拼法。
最初發現的傷齒龍牙齒化石,由約瑟夫·萊迪繪製
傷齒龍的牙齒起初被萊迪分類於蜥蜴亞目。但在1901年,法蘭茲·諾普喬(Franz Nopcsa)將傷齒龍重新歸類於斑龍科;斑龍科在歷史上長時間為大部分肉食性恐龍的集中地。在1924年,吉爾摩爾提出這些牙齒應屬於草食性厚頭龍類的劍角龍,而劍角龍其實是傷齒龍的一個次異名;傷齒龍科牙齒與草食性恐龍牙齒之間的相似處,讓許多古生物學家認為這些動物是雜食性動物。在1945年,查爾斯·斯騰伯格(Charles Mortram Sternberg)否決傷齒龍屬於厚頭龍類的可能性,因為這些牙齒與肉食性恐龍的牙齒有較多相似處。
傷齒龍的第一個標本並不是牙齒,而是斯騰伯格在1932年所命名的細爪龍化石,細爪龍的化石被發現於亞伯達省,由一個腳部、手部的碎片、以及一些尾椎所構成。這些化石的明顯特徵是第二腳趾上的加大趾爪,這被認為是恐爪龍下目的特徵。斯騰伯格起初將細爪龍分類於虛骨龍科。在1951年,斯騰伯格假設因為細爪龍有非常特別的足部,而傷齒龍有一樣獨特的牙齒,它們可能為近親。不幸的是,在當時並沒有足夠的標本可供比較,來驗證這個假設。
傷齒龍的重建模型,位於倫敦自然歷史博物館
在1969年,戴爾·羅素(Dale Russell)敘述了一個更完整的細爪龍骨骸;細爪龍演化成為類似人類的虛構生物“類恐龍人”的科學理論基礎,來自於這個研究。細爪龍在1980年代成為一個著名的獸腳類恐龍,而它們的腦部與腳部被敘述地更詳盡。在1987年,菲力·柯爾(Phil Currie)重新審視已知的傷齒龍科化石,他將細爪龍重新分類為美麗傷齒龍的一個異名。這次改變被其他古生物學家廣泛地接受,因此在最近的科學文獻中,所有過去被稱為細爪龍的標本,現在都被改稱為傷齒龍。
傷齒龍是種小型恐龍,身長約2米,高度為1米,重達60公斤。傷齒龍擁有非常修長的四肢,顯示它們可以快速奔跑。傷齒龍擁有長手臂,可以像鳥類一樣往後折起,而手部擁有可作出相對動作的拇指。它們的第二腳趾上擁有大型、可縮回的鐮刀狀趾爪,這些趾爪在奔跑時可能會抬起。
因為這些特徵,科學家們認為傷齒龍屬於手盜龍類。傷齒龍擁有大型眼睛,顯示可能有夜間行動,可能以夜間行動的哺乳動物為食;而且眼睛的方向稍微往前,使傷齒龍擁有深度知覺。事實上傷齒龍的眼睛比其他大部分恐龍還要朝向前方,顯示傷齒龍有比其他恐龍更好的深度知覺。它們輕型頭顱骨的腦囊類似某些外表類似鴕鳥的恐龍的腦囊。與身體相比,傷齒龍的腦袋是恐龍中最大的。因此傷齒龍被認為是最有智能的恐龍之一。曾經在蛋巢中發現過傷齒龍的恐龍蛋。
傷齒龍的化石被發現於蒙大拿州的朱迪斯河組與海爾河組及雙麥迪遜組、亞伯達省的馬蹄峽谷組、以及阿拉斯加州的北坡(North Slope)地區。還有一些證據顯示傷齒龍較常生活在寒冷的地區,它們似乎在北部地區特別繁盛,以及較寒冷的時期,例如早馬斯特里赫特階。
這些化石來自於相隔數千公里遠的地點,而且是在各大陸分隔數百萬年之後出現,似乎不可能代表傷齒龍屬的同一個種。然而,需要進一步的研究與更多化石才能理解傷齒龍到底有幾個種。
1978至1984年間,約翰·霍納(John Horner)和同事鮑勃·馬凱拉(Bob Makela)在蒙大拿山區進行挖掘,發現了三處恐龍巢穴遺跡,包括一窩排列整齊的傷齒龍(Troodon)蛋。傷齒龍把卵產在剛乾涸的湖底或沼澤地的濕潤泥土裡,靠輸卵管向下蠕動的力量能輕鬆地把它們深深插入泥土中。而生活在中國的白堊紀傷齒龍則是選擇水邊的沙土地作為產卵地點,它們先用爪子在地上刨出一個坑,然後蹲坐下來使身子成直立或半直立狀態,並把蛋產入蛋坑裡鬆軟的沙土中。之後再用沙土小心地地這些蛋埋起來。它們還會孵蛋。
一個傷齒龍的蛋巢,在落磯山博物館展出中
在2002年,Varriccho等人敘述了八個在蒙大拿州的雙麥迪遜組發現的傷齒龍蛋巢。這些蛋巢目前存放在落磯山博物館。第一個蛋巢是由傑克·霍納(Jack Horner)在1983年發現。霍納在隔年發現相關的骨頭,並在蛋巢附近發現了棱齒龍類奔山龍的部分骨骸,霍納因此將這些蛋敘述為奔山龍的蛋。在1996年,霍納與大衛·威顯穆沛(David B. Weishampel)重新檢驗蛋中的胚胎,認為這些蛋屬於傷齒龍,而非奔山龍。
在1997年,Varricchio等人敘述了另一個成年傷齒龍的部分骨骸,與其懐中的至少五顆蛋,這隻恐龍可能正在孵蛋。Varricchio等人也敘述了上述其中一個蛋巢。這個蛋巢呈碟狀,內側直徑約100厘米,巢壁將蛋圍在內側。其他更完整的蛋巢則有16到24顆蛋。這些蛋的外形為延長的淚滴狀,下端尖,下半部插入到沉積層內,上半部傾斜,往中心聚攏。沒有證據顯示蛋巢內有植物材料。
Varricchio等人從這些蛋巢,提出了數個傷齒龍繁衍能力特徵。傷齒龍的繁衍模式似乎介於鱷魚與鳥類之間,如同種系發生學的默認結果。這些但以成對方式排列,顯示傷齒龍可能具有兩個產道,類似鱷魚,而非僅有一個的鳥類。鱷魚產的蛋數量較多,蛋與成年體的身體相比較小;鳥類的蛋數量較少,與身體相比較大。傷齒龍的體積比則位居中間,蛋的重量約0.5公斤,而成年體的重量為50公斤。這個比例是同等體積爬蟲類的10倍,但其中兩顆蛋的重量約1.1公斤,比例接近同等體積的鳥類。Varricchio等人也發現多次產卵的證據,一個成年傷齒龍可能在每一、兩天產兩顆蛋。其中一個蛋巢有22個蛋,而另一蛋巢中的胎胚發展程度接近,顯示這些胚胎孵化模式接近。從這些胚胎的發展模式顯示,傷齒龍可能剛孵化不久就可以行動,甚至是剛孵化就可以。研究人員估計成年傷齒龍從產卵、孵化蛋、到出生,約化45到65天。研究人員發現蛋巢內沒有幼年傷齒龍活動的證據,顯示它們可能剛孵化就離開蛋巢,如同鱷魚與冢雉。
在2008年,Varricchio等人檢驗編號MOR 748標本的骨組織,發現該標本的骨頭缺乏吸收骨髓骨的跡象,顯示該個體是個正在產卵中的個體。這群科學家也測量傷齒龍蛋巢中的蛋總體積,與成年傷齒龍的體積比例;將所測量的比例與現代鳥類、鱷魚相比時,發現傷齒龍的數值與雄性會孵化蛋的鳥類相近。根據此結果,他們推測雌性傷齒龍沒有孵化蛋的習性,而是由雄性傷齒龍孵化蛋;另外,他們也推測當時的手盜龍類與原始鳥類也擁有相同的習性。
傷齒龍科恐龍的化石是第一群被敘述的恐龍化石之一。最初,在1856年約瑟夫·萊迪(JosephLeidy)將這些動物分類於蜥蜴亞目;但在1924年,查爾斯·惠特尼·吉爾摩(CharlesW.Gilmore)將他們分類於恐龍歸目的鳥臀目。直到1945年,查爾斯·斯騰伯格(CharlesMortramSternberg)將傷齒龍科鑒定、分分類於獸亞亞目。1969年以來,傷齒龍科與馳龍科共同屬於恐爪龍下目演化支。但在1994年,托馬斯·霍爾特(ThomasR.HoltzJr)根據加大的腦部、長而低的上頜孔等特徵,建立了Bullatosauria,包含了似鳥龍下目與傷齒龍科。傷齒龍科的骨盆也顯示他們較馳龍科原始。在中國新發現的原始傷齒龍科化石,例如中國列龍與寐龍,顯示出傷齒龍科、馳龍科、以及原始鳥類始祖鳥之間的相似處;而現在的大多數古生物學家,包括霍爾特在內,都認為傷齒龍科較接近鳥類,而離似鳥龍下目較遠,使得Bullatosauria分類遭到廢棄。由TheropodWorkingGroup成員所進行的密集獸腳亞目研究,顯示基礎馳龍科恐龍、傷齒龍科、以及始祖鳥之間有非常相似的地方。這幾個分類形成近鳥類演化支,該演化支由Novas與Pol所提出。Hwang等人所提出的演化樹,顯示始祖鳥代表近鳥類的一個原始分支,而馳龍科與與齒龍科較為先進。這個理論提升了恐爪龍類演化出空氣動力行為的可能性。