電感受器
電感受器
電感受器(electroreceptor)是能夠接受外界微弱電流而發放傳入神經衝動的感受器。僅在某些魚類(如電魚和軟骨魚綱魚類)中存在。例如鯊及鰩皮膚深部的洛倫齊尼氏壺腹就是一種電感受器,它對電流特別敏感。狗鯊和射鰩的電感受器能對5×10^-11安培的微弱電流發生反應。
電感受器通常是由位於體表的側線器官變態而成,因此對機械刺激也有反應。電感受器管腔內充滿高電導的腖膠樣物質。感受器具有外界電場0.1微伏/厘米就有充分反應的能力。據說,以此可以探知食餌動物的肌肉動作電位而用於捕食。硬骨魚類的鯰鰻也有同樣的器官。另一方面,裸背鰻科(Gymnotidae)、長頜魚科(Mormyridae)、裸臀魚科(Gymnarchidae)的某種小形淡水魚稱為弱電魚,具有自身的發電器作為脈衝放電,在身體周圍形成電場,通過體表的感覺器來檢測導體和非導體接近時引起電場的擾亂狀態。這種感受器系側線器官的變形而成,與勞倫齊尼器官略有不同,不在皮膚開口處,而是隱於皮下。放電頻率隨種類而異,但電敏感性總在0.1—100微伏/厘米的範圍內。
電感受器是多種多樣的,但按其結構與功能特點可分為緊張性電感受器(ampullary organ)和位相性電感受器(tuberous organ)兩種基本類型。
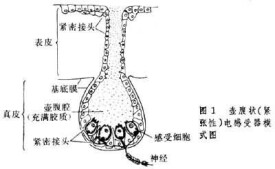
緊張性電感受器
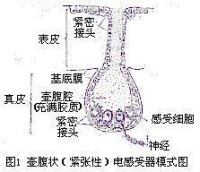
緊張性電感受器模式圖
位相性電感受器
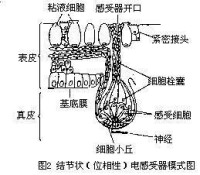
位相性電感受器模式圖
當電流作用於電感受器細胞時,由於這些細胞的細胞膜的不同部位具有不同的電阻,細胞上部的細胞膜電阻小,電流易於通過,但其底膜(細胞的基底部)電阻大,這樣就限制了外在電流從外向內通過。密集的電流使底膜迅速去極化,並導致突觸遞質的釋放速度大大超過自發性釋放的速度,因而使支配感受器細胞的神經纖維發放傳入衝動的頻率增加。當外界電流通過魚體以後,這些細胞的底膜出現超極化,使遞質的釋放速度小於自發性釋放的速度,這時神經纖維發放衝動的頻率也隨之相應減少。這樣,傳入神經衝動發放頻率的高低,可以由電流通過感受器細胞來決定。因此,具有電感受器的魚類能夠覺察外界微弱電流的存在,並對它發生反應。
電感受器所感受水中的電源,是來自電魚的放電器官或其他非電魚的神經肌肉活動時所產生的微弱電活動。例如,非洲電魚能連續地發放頻率約為300~400次/秒的電脈衝。當每次發放電脈衝時,尾部的尖端較頭部為負。這樣,電流就通過水而到達頭部並形成電場。電場的結構決定於環境的導電性,假若其間有個導電性比水高或比水低的物體,則電場的結構發生改變。導電性高的物體使電流線集中,導電性低的物體會使電流線分散。電感受器能夠感受電場的這些變化。因此,電魚利用自身的電器官放電和電感受器感受電流的特性,可以探測環境和確定外在物體的位置。有些魚類沒有電器官,不發放電信號,但卻具有電感受器,能夠感受其他動物的肌肉(例如,呼吸肌和游泳肌)所產生的微弱電流。鯊和鰩可以通過這種方式來尋找其他魚類,即使那些魚停止遊動或潛在泥沙中不動也逃脫不了。例如,鯊可以找到埋藏在水族箱的沙底里的鰈,假如鯊從離鰈不到15厘米的地方經過,它會立即轉向埋藏的鰈,將之吸起獵獲。