共找到2條詞條名為神經遞質的結果 展開
- 神經遞質
- 神經介質
神經遞質
神經遞質
神經遞質(neurotransmitter)是神經元之間或神經元與效應器細胞如肌肉細胞、腺體細胞等之間傳遞信息的化學物質。根據神經遞質的化學組成特點,主要有膽鹼類(乙醯膽鹼,acetylcholineAch)、單胺類(去甲腎上腺素、多巴胺和5-羥色胺)、氨基酸類(興奮性遞質如谷氨酸和天冬氨酸;抑制性遞質如γ氨基丁酸、甘氨酸和牛磺酸)和神經肽類等。在神經元的信息傳遞過程中,當一個神經元受到來自環境或其他神經元的信號刺激時,儲存在突觸前囊泡內的遞質可向突觸間隙釋放,作用於突觸后膜相應受體,將遞質信號傳遞給下一個神經元。神經遞質主要以旁分泌方式傳遞信號,因此速度快、準確性高。遞質信號的終止可依賴於突觸間隙或后膜上相應的水解酶分解破壞,或者被突觸前膜特異性遞質轉運體重攝取。
在中樞神經系統(CNS)中,突觸傳遞最重要的方式是神經化學傳遞。神經遞質的作用可通過兩個途徑中止:一是再回收抑制,即通過突觸前載體的作用將突觸間隙中多餘的神經遞質回收至突觸前神經元並貯存於囊泡;另一途徑是酶解,如以多巴胺(DA)為例,它經由位於線粒體的單胺氧化酶(MAO)和位於細胞質的兒茶酚胺鄰位甲基轉移酶(COMT)的作用被代謝和失活。
遞質的代謝包括合成、儲存、釋放和滅活四個環節。
乙醯膽鹼(Ach)的合成主要是在膽鹼能神經末梢內進行。由膽鹼和乙醯輔酶A在膽鹼乙醯化酶的催化下合成乙醯膽鹼,然後轉移到囊泡儲存:當神經衝動到達神經末梢時,囊泡膜與突觸前膜相融合將乙醯膽鹼釋放入突觸間隙,激動突觸后膜上相應受體,引起一系列生理效應。同時,乙醯膽鹼由神經末梢部位的膽鹼酯酶(ChE)水解為膽鹼和乙酸而滅活。部分膽鹼再一次被膽鹼能神經末梢攝取,又參與合成新的乙醯膽鹼。
去甲腎上腺素(NA)的合成主要在去甲腎上腺素能神經末梢內進行。由腎上腺素能神經末梢的胞漿攝取血中酪氨酸,在酪氨酸羥化酶和脫羧酶催化下轉變成多巴胺,再經多巴胺β-羥化酶催化合成去甲腎上腺素,儲存於囊泡中。當神經衝動到達神經末梢時,囊泡向突觸前膜靠近,以胞裂外排的方式釋放去甲腎上腺素到突觸間隙,激動突觸后膜上相應的受體產生一系列生理效應。
釋放后的去甲腎上腺素,大部分(75%~90%)被突觸前膜再攝入神經末梢內,轉入囊泡內儲存,僅有小部分被突觸間隙的兒茶酚氧位甲基轉移酶(COMT)和單胺氧化酶(MAO)滅活。
作為神經遞質應具備以下條件:
①在突觸前神經元內具有能合成遞質的物質及酶系統;
②遞質貯存於突觸小泡內,不被胞漿內其他酶所破壞,在神經衝動到達時,能被釋放進入突觸間隙;
③遞質通過突觸間隙,能夠作用於突觸后膜的特殊受體,產生突觸后電位;
神經遞質
④遞質能迅速失活;
⑤能人工地把該物質直接作用於突觸后膜,產生與突觸前膜釋放該遞質相同的生理效應;
⑥其作用能被特異性藥物阻斷或加強。
腦內神經遞質分為四類,即生物原胺類、氨基酸類、肽類、其它類。生物原胺類神經遞質是最先發現的一類,包括:多巴胺(DA)、去甲腎上腺素(NE)、腎上腺素(E)、5-羥色胺(5-HT)也稱(血清素)。氨基酸類神經遞質包括:γ-氨基丁酸(GABA)、甘氨酸、谷氨酸、組胺、乙醯膽鹼(Ach)。肽類神經遞質分為:內源性阿片肽、P物質、神經加壓素、膽囊收縮素(CCK)、生長抑素、血管加壓素和縮宮素、神經肽y。其它神經遞質分為:核苷酸類、花生酸鹼、阿南德醯胺、sigma受體(σ受體)。其它類:一氧化氮就被普遍認為是神經遞質,它不以胞吐的方式釋放,而是憑藉其溶脂性穿過細胞膜,通過化學反應發揮作用並滅活。在突觸可塑性變化、長時程增強效應中起到逆行信使的作用。
重要的神經遞質和調質有:
①乙醯膽鹼。最早被鑒定的遞質。脊椎動物骨骼肌神經肌肉接頭、 某些低等動物如軟體、環節和扁形動物等的運動肌接頭等,都是以乙醯膽鹼為興奮性遞質。脊椎動物副交感神經與效應器之間的遞質也是乙醯膽鹼,但有的是興奮性的(如在消化道),有的是抑制性的(如在心肌)。中國生理學家張錫鈞和J.H.加德姆(1932)所開發的以蛙腹直肌標本定量測定乙醯膽鹼的方法,對乙醯膽鹼的研究起了重要作用,至今仍有應用價值。
某些低等動物如軟體、環節和扁形動物等的運動肌接頭等,都是以乙醯膽鹼為興奮性遞質。脊椎動物副交感神經與效應器之間的遞質也是乙醯膽鹼,但有的是興奮性的(如在消化道),有的是抑制性的(如在心肌)。中國生理學家張錫鈞和J.H.加德姆(1932)所開發的以蛙腹直肌標本定量測定乙醯膽鹼的方法,對乙醯膽鹼的研究起了重要作用,至今仍有應用價值。
神經遞質
②兒茶酚胺。包括去甲腎上腺素(NE)、腎上腺素(E)和多巴胺(DA)。交感神經節細胞與效應器之間的接頭是以去甲腎上腺素為遞質。
③5-羥色胺(5-HT)。5-羥色胺神經元主要集中在腦橋的中縫核群中,一般是抑制性的,但也有興奮性的。中國一些學者的研究表明,在針刺鎮痛中5-羥色胺起著重要作用。
④氨基酸遞質。被確定為遞質的有谷氨酸(Glu)、γ-氨基丁酸(GABA)和甘氨酸(Gly)。谷氨酸是甲殼類神經肌肉接頭的遞質。γ氨基丁酸首先是在螯蝦螯肢開肌與抑制性神經纖維所形成的接頭處發現的遞質。後來證明γ-氨基丁酸也是中樞的抑制遞質。以甘氨酸為遞質的突觸主要分佈在脊髓中,也是抑制性遞質。
突觸傳遞是通過突觸前膜釋放化學遞質來完成的(非突觸性化學傳遞的情況也是如此)。一個化學物質被確認為神經遞質,應符合以下條件:
①在突觸前神經元內具有全盛遞質的前體物質和合成酶系,能夠合成這一遞質;
②遞質貯存於突觸小泡以防止被胞漿內其它酶系所破壞,當興奮衝動抵達神經末梢時,小泡內遞質能釋放入突觸間隙;
③遞質通過突觸間隙作用於突觸后膜的特殊受體,發揮其生理作用,用電生理微電泳方法將遞質離子施加到神經元或效應細胞旁,以模擬遞質釋放過程能引致相同的生理效應;
④存在使這一遞質失活的酶或其他環節(攝取回收);
⑤用遞質擬似劑或受體阻斷劑能加強或阻斷這一遞質的突觸傳遞作用。在神經系統內存在許多化學物質,但不一定都是神經遞質,只有符合或基本上符合以上條件的化學物質才能認為它是神經遞質。關於神經遞質,首先是在外周迷走神經對心臟抑制作用的環節上發現的。
膽鹼能
乙醯膽鹼在蛙心灌注實驗中觀察到,刺激迷走神經時蛙心活動受到抑制,如將灌流液轉移到另一蛙心製備中去,也可引致后一個蛙心的抑制。顯然在迷走神經興奮時,有化學物質釋放出來,從而導致心臟活動的抑制。後來證明這一化學物質是乙醯膽鹼,乙醯膽鹼是迷走神經釋放的遞質。以後在許多其他器官中(例如胃腸、膀胱、頜下腺等),刺激其副交感神經也可在灌注液中找到乙醯膽鹼。由此認為,副交感神經節后纖維都是釋放乙醯膽鹼作為遞質的。釋放乙醯膽鹼作為遞質的神經纖維,稱為膽鹼能纖維。
自主神經系統神經末梢的化學傳遞
人進行了上頸交感神經節的灌流,見到刺激節前纖維可以灌流液中獲得乙醯膽鹼,所以節前纖維的遞質也是乙醯膽鹼。現已明確軀體運動纖維也是膽鹼能纖維。節前纖維和運動神經纖維所釋放的乙醯膽鹼的作用,與菸鹼樣作用(N樣作用);而副交感神經節后纖維所釋放的乙醯膽鹼的作用,也毒蕈鹼的藥理作用相同,稱為毒蕈鹼樣作用(M樣作用)。
去甲腎上腺素能
去甲腎上腺素交感神經節后纖維的遞質比較複雜。本世紀初,有人見到腎上腺素對效應器的廣泛作用與交感神經的作用極為相似,因此設想交感神經可能是通過末梢釋放腎上腺素而對效應器起作用的。後來,在貓的實驗中觀察到,刺激支配尾巴的交感神經可以引致尾巴上毛的豎立和血管收縮,同時該動物的去神經支配的心臟活動加速;如果將自尾巴迴流的靜脈結紮,再刺激這一交感神經就只能引致尾巴上毛的豎立和血管收縮,卻不能引致心臟活動的加速。由此設想,支配尾巴的交感神經末梢能釋放一種化學物質,由靜脈迴流於心臟,這種物質在當時稱為交感素。交感素比乙醯膽鹼的性質穩定,當有大量釋放時不易破壞,在一般情況下有可能經血液循環作用於較為遠隔的效應器官。後來,在刺激支配其他器官的交感神經時,均證明靜脈血中出現交感素。曾有人指出,交感素是去甲腎上腺素和腎上腺素的混合物,而主要是去甲腎上腺素。現已明確,在高等動物中由交感神經節后纖維釋放的遞質僅是去甲腎腺上素,而不含腎上腺素;因為在神經末梢只能合成去甲腎上腺素,而不能進一步合成腎上腺素,由於末梢中不含合成腎上腺素所必需的苯乙醇胺氮位甲基移位酶。釋放去甲腎上腺素作為遞質的神經纖維,稱為腎上腺素能纖維。但是,不是所有的交感神經節后纖維都是腎上腺素能纖維,像支配汗腺的交感神經和骨骼肌的交感舒血管纖維卻是膽鹼能纖維。
嘌呤類和肽類遞質
嘌呤類和肽類遞質自主神經的節后纖維除膽鹼能和腎上腺素能纖維外,還有第三類纖維。第三類纖維末梢釋放的遞質是嘌呤類和肽類化學物質。有人在實驗中觀察到,刺激這類神經時實驗標本灌流液中可以找到三磷酸腺苷及其分解產物;而三磷酸腺苷對有腸肌的作用與這類神經的作用極相似,兩者均可引致腸肌的舒張和腸肌細胞電位的超極化。因此認為這類神經末梢釋放的遞質是三磷酸腺苷,是一種腺嘌呤化合物。但也有人認為這類神經釋放的遞質是肽類化合物,因為免疫細胞化學的研究證實自主神經某些纖維末梢的大顆粒囊泡中含有血管活性腸肽,刺激迷走神經時能引致血管活性腸肽的釋放。血管活性腸肽能使胃腸平滑肌舒張,胃的容受性舒張可能就是由於迷走神經節后纖維釋放血管活性腸肽遞質而實現的。第三類纖維是非膽鹼能和非腎上腺素能纖維,主要存在於胃腸,其神經元細胞體位於壁內神經叢中;在胃腸上部它接受副交感神經節前纖維的支配。
閏紹細胞(Renshaw cell)是脊髓前角內的一種神經元,它接受前角運動神經元軸突側支的支配,它的活動轉而反饋抑制前角運動神經元的活動。前角運動神經元支配骨骼肌的接頭處遞質為乙醯膽鹼,則其軸突側支與閏紿細胞發生突觸聯繫,也必定釋放乙醯膽鹼作為遞質。用電生理微電泳法將乙醯膽鹼作用於閏紹細胞,確能引致其放電;用N型受體阻斷劑后,乙醯膽鹼的興奮作用即被阻斷,說明這一突觸聯繫的乙醯膽鹼作用與神經肌接頭處一樣都是N樣作用.
脊髓前角運動神經元與閏紹細胞的反饋聯繫
位於丘腦後部腹側的特異感覺投射神經元是膽鹼能神經元,它們和相應的皮層感覺區神經元形成的突觸是以乙醯膽鹼為遞質的。例如,刺激視神經時,枕葉皮層17區等處的乙醯膽鹼釋放增多。
腦幹網狀結構上行激動系統(參見第三節)的各個環節似乎都存在乙醯膽鹼遞質。例如,腦幹腦狀結構內某些神經元對乙醯膽鹼敏感;刺激中腦網狀結構使腦電出現快波時,皮層的乙醯膽鹼釋放明明顯增加;用組織化學法顯示腦幹網狀結構的乙醯膽鹼上行通路,發現其與腦幹網狀結構上行激動系統通路有相似之外。
尾核含有豐富的乙醯膽鹼、膽鹼乙醯移位酶和膽鹼酯酶,尾核內有較多的神經元對乙醯膽鹼敏感,殼核與蒼白球內某些神經元也對乙醯膽鹼敏感。由此看來,紋狀體內存在乙醯膽鹼遞質系統。
綜上所述,乙醯膽鹼肯定是中樞的遞質,而且分佈比較廣泛。
單胺類遞質是指多巴胺、去甲腎上腺素和5-羥色胺。由於動物實驗中採用了熒光組織化學方法,對中樞內單胺類遞質系統了解得比較清楚。
單胺類遞質的通徑 多巴胺遞質系統主要包括三部位:黑質-紋狀體部分、中腦邊緣系統部分和結節、漏斗部分。黑質-紋狀體部分的多巴胺能神經元位於中腦黑質,其神經纖維投射到紋狀體。腦內的多巴胺主要由黑質製造,沿黑質-紋狀體投射系統分佈,在紋狀體貯存(其中以尾核含量最多)。破壞黑質或切斷黑質-紋狀體束,紋狀體中多巴胺的含量即降低。用電生理微電泳法將多巴胺作用於紋狀體神經元,主要起抑制反應。中腦位於邊緣部分的多巴胺能神經元位於中腦腳間核頭端的背側部位,其神經纖維投射到邊緣前腦。結節-漏斗部分的多巴胺能神經元位於下丘腦弓狀核,其神經纖維投射到正中隆起。
多巴胺遞質系統主要包括三部位:黑質-紋狀體部分、中腦邊緣系統部分和結節、漏斗部分。黑質-紋狀體部分的多巴胺能神經元位於中腦黑質,其神經纖維投射到紋狀體。腦內的多巴胺主要由黑質製造,沿黑質-紋狀體投射系統分佈,在紋狀體貯存(其中以尾核含量最多)。破壞黑質或切斷黑質-紋狀體束,紋狀體中多巴胺的含量即降低。用電生理微電泳法將多巴胺作用於紋狀體神經元,主要起抑制反應。中腦位於邊緣部分的多巴胺能神經元位於中腦腳間核頭端的背側部位,其神經纖維投射到邊緣前腦。結節-漏斗部分的多巴胺能神經元位於下丘腦弓狀核,其神經纖維投射到正中隆起。
神經遞質
去甲腎上腺素系統比較集中,極大多數的去甲腎上腺素能神經元位於低位腦幹,尤其是中腦網狀結構、腦橋的藍斑以及延髓網狀結構的腹外側部分。按其纖維投射途徑的不同,可分為三部分:上行部分、下行部分和支配低位腦幹部分。上行部分的纖維投射到大腦皮層,邊緣前腦和下丘腦。下行部分的纖維下達脊髓背角的膠質區、側角和前角。支配低位腦幹部分的纖維,分佈在低位腦幹內部。
5-羥色胺遞質系統也比較集中,其神經元主要位於低位腦幹近中線區的中縫核內。按其纖維投射途徑的不同,也可分為三部分:上行部分、下行部分和支配低位腦幹部分。上行部分的神經元位於中縫核上部,其神經纖維投射到紋狀體、丘腦、下丘腦、邊緣前腦和大腦皮層。腦內5-羥色胺主要來自中縫核上部,破壞中縫核上部可使腦內5-羥色胺含量明顯降低。下行部分的神經元位於中縫核下部,其神經纖維下達脊髓背角的膠質區、側角和前角。支配低位腦幹部分的纖維,分佈在低位腦幹內部。
氨基酸類 現快明確存在氨基酸類遞質,例如谷氨酸、門冬氨酸、甘氨酸和γ-氨基丁酸。
在腦脊髓內谷氨酸含量很多,分佈很廣,但相對來看,大腦半球和脊髓背側部分含量較高。用電生物微電泳法將谷氨酸作用於皮層神經元和脊髓運動神經地,可引致突觸后膜出現類似興奮性突觸后電位的反應,並可導致神經元放電。由此設想,谷氨酸可能是感覺傳入神經纖維(粗纖維類)和大腦皮層內的興奮型遞質。
用電生理微電泳法將甘氨酸作用於脊髓運動神經元,可引致突觸后膜出現類似抑制性突觸后電位的反應。閏紹細胞軸突末梢釋放的遞質就是甘氨酸,它對運動神經元起抑制作用。
γ-氨基丁酸在大腦皮層的淺層和小腦皮層的浦肯野細胞層含量較高。用電生理微電泳法將γ-氨基丁酸作用於大腦皮層神經元和前庭外側核神經元(直接受小腦皮層浦肯野細胞支配),可引致突觸后膜超極化。由此設想,γ-氨基丁酸可能是大腦皮層部分神經元和小腦皮層浦肯野細胞的抑制性遞質。此外,紋狀體-黑質的纖維,也是釋放γ-氨基西酸遞質的。
上述的抑制是突觸后膜發生超極化而發生的,因此是突觸后抑制。所以甘氨酸和γ-氨基丁酸均是突觸后抑制的遞質。已知,γ-氨基丁酸也是突觸前抑制的遞質;當γ-氨基丁酸作用於軸突末梢時可引致末梢支極化,使末梢在衝動抵達時遞質釋放量減少,從而產生抑制效應(參見第二節)。γ-氨基丁酸對細胞體膜產生超極化,而對末梢軸突膜卻產生去極化,其機制尚不完全清楚。有人認為,γ-氨基丁酸的作用是使膜對CI-的通透性增升高;在細胞體膜對CI-的通透性升高時,由於細胞外CI-濃度比細胞內CI-濃度高,CI-由細胞外進入細胞內,因此產生超極化;在末梢軸突膜對CI-通透性升高時,由於軸漿內CI-濃度比軸突外CI-高,CI-由軸突內流向軸突外,因此產生去極化。所以γ-氨基丁酸的作用是使CI-通透性升高,造成超極化還是去極化,取決於細胞內外CI-的濃度差。
早已知道神經元能分泌肽類化學物質,例如視上核和室旁核神經元分泌升壓素(九肽)和催產素(九肽);下丘腦內其他肽能神經元能分泌多種調節腺垂體活動的多肽,如促甲狀腺釋放激素(TRH,三肽)、促性腺素釋放激素(GnRH,十肽)、生長抑素(GHRIH,十四肽)等。由於這些肽類物質在分泌后,要通過血液循環才能作用於效應細胞,因此稱為神經激素。但現已知,這些肽類物質可能還是神經遞質。例如,室旁核有向腦幹和脊髓投射的纖維,具有調節交感和副交感神經活動的作用(其遞質為催產素),並能抑制痛覺(其遞質為升壓素)。在下丘腦以外腦區存在TRH和相應的受體,TRH能直接影響神經元的放電活動,提示TRH可能是神經遞質。
腦內具有嗎啡樣活性的多肽,稱為阿片樣肽。阿片樣肽包括β-內啡肽、腦啡肽和強啡肽三類。腦啡肽是五肽化合物,有甲硫氨酸腦啡肽(M-ENK)和亮氨酸腦啡肽(L-ENK)兩種。腦啡肽與阿片受體常相伴而存在,微電泳啡肽可命名大腦皮層、紋狀體和中腦導水管周圍灰質神經元的放電受到抑制。腦啡肽在脊髓背角膠質區濃度很高,它可能是調節痛覺纖維傳入活動的神經遞質。
腦內還有胃腸肽存在,例如膽囊收縮素(CCK)、促胰液素、胃泌素、胃動素、血管活性腸肽、胰高血糖素等。CCK有抑制攝食行為的作用。許多膽鹼能神經元中含有血管活性腸肽,它可能具有加強乙醯膽鹼作用的功能。此外,腦內還有其他肽類物質,例如P物質、神經降壓素、血管緊張素Ⅱ等。P物質是十一肽,它可能是第一級感覺神經元(屬於細纖維類)釋放的興奮性遞質,與痛覺傳入活動有關。神經降壓素在邊緣系統中存在。血管緊張素Ⅱ的主要作用可能在於調節單受類纖維的遞質釋放。
其他可能的遞質研究指出,一氧化氮具有許多神經遞質的特徵。某些神經元含有一氧化氮合成酶,該酶能使精氨酸生成一氧化氮。生成的一氧化氮從一個神經元彌散到另一神經元中,而後作用於鳥苷酸環化酶並提高其活力,從而發揮出生理作用。因此,一氧化氮是一個神經元間信息溝通的傳遞物質,但與一般遞質有區別:
①它不貯存於突觸小泡中;
②它的釋放不依賴於出胞作用,而是通過彌散;
③它不作用於靶細胞膜上的受體蛋白,而是作用於鳥苷酸環化酶。一氧化氮與突觸活動的可塑性可能有關,因為用一氧化氮合成酶抑製劑后,海馬的第時程增強效應被完全阻斷(參見第六節中“學習和記憶的機制”)。此外,組織胺也可能是腦內的神經遞質。
是指神經末梢釋放的特殊化學物質,它能作用於支配的神經元或效應細胞膜上的受體,從而完成信息傳遞功能。
是指神經元產生的另一類化學物質,它能調節信息傳遞的效率,增強或削弱遞質的效應。
但是也有人把遞質概念規定得非常嚴格,認為只有作用於膜受體后導致離子通道開放從而產生興奮或抑制的化學物質才能稱為遞質;其他一些作用於膜受體后通過第二信使轉而改變膜的興奮性或其它遞質釋放的化學物質,均應稱為調質。根據后一種觀點,遞質為數不多,氨基酸類物質是遞質,神經肌接頭部位釋放的乙醯膽鹼也是遞質,而肽類物質一般均屬於調質。但是一般來說,遞質與調質無明確劃分的界限,調質是從遞質中派生出來的概念,不少情況下遞質包含調質;前文就沒有把兩者嚴格區分開來,統稱為遞質。
長期來認為,一個神經元內只存在一種遞質,其全部神經末梢均釋放同一種遞質。這一原則稱為戴爾原則(Dale’s principle)。通過免疫組織化學方法觀察到,一個神經元內可存在兩種或兩種以上遞質(包括調質),因此認為戴爾原則並不正確。但是戴爾的原先觀點認為,一個神經元的全部神經末梢均釋放相同的遞質;他並沒有限定一個神經元只能含一種遞質。因此,戴爾的觀點還是對的,而戴爾原則則是需要修改的。
在無脊椎動物的神經元中,觀察到多巴胺和5-羥色胺遞質可以共存。在高等動物的交感神經節神經節發育過程中,去甲腎上腺素和乙醯膽鹼可以共存。此外,在大鼠延髓的神經元中觀察到5-羥色胺和P物質共存;在上頸交感神經節中神經元中觀察到去甲腎上腺素和腦啡肽共存。有人認為肽類遞質可能都是與其他遞質共存的。遞質共存的生理意義,目前尚未清楚了解;可能兩種遞質在同時釋放後起著不同的生理作用,有利於發揮突觸傳遞作用。
乙醯膽鹼是由膽鹼和乙醯輔酶A在膽鹼乙醯移位酶(膽鹼乙醯化酶)的催化作用下合成的。由於該酶存在於胞漿中,因此乙醯膽鹼在胞漿中合成,合成後由小泡攝取並貯存起來。去甲腎上腺素的合成以酪氨酸為原料,首先在酪氨酸羥化酶的催化作用下合成多巴,再在多巴脫羧酶(氨基酸脫竣酶)作用下合成多巴胺(兒茶酚乙胺),這二步是在胞漿中進行的;然後多巴胺被攝取入小泡,在小泡中由多巴胺β羥化酶催化進一步合成去甲腎上腺素,並貯存於小泡內。多巴胺的合成與去甲腎上腺素前二步是完全一樣的,只是在多巴胺進入小泡后不再合成去甲腎上腺素而已,因為貯存多巴胺的小泡內不含多巴胺β羥化酶。5-羥色胺的合成以色氨酸為原料,首先在色氨酸羥化酶作用下合成5-羥色氨酸,再在5-羥色胺酸脫羧酶(氨基酸脫羧酶)作用下將5-羥色氨酸合成5-羥色胺,這二步是在胞漿中進行的;然後5-羥色胺被攝取入小泡,並貯存於小泡內。γ-氨基丁酸是谷氨酸在谷氨酸脫羧催化作用下合成的。肽類遞質的合成與其他肽類激素的合成完全一樣,它是由基因調控的,並在核糖體上通過翻譯而合成的。
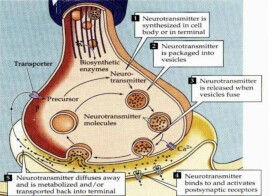
當神經衝動抵達末梢時,末梢產生動作電位和離子轉移Ca2+由膜外進入膜內,使一定數量的小泡與突觸前膜緊貼融合起來,然後小泡與突觸前膜粘合處出現破裂口,小泡內遞質和其他內容物就釋放到突觸間隙內。突觸前膜釋放遞質的過程,稱為出胞(exocytosis)或胞裂外排。在這一過程中,Ca2+的轉移很重要。如果減少細胞外Ca2+濃度,則遞質釋放就受到抑制;而增加細胞外Ca2+的濃度則遞質釋放增加。這一事實說明,Ca2+由膜外進入膜內的數量多少,直接關係到遞質的釋放量;Ca2+是小泡膜與突觸前膜緊貼融合的必要因素。一般認為,Ca2+可能有兩方面的作用:
①降低軸漿的粘度,有利於小泡的移動;
②消除突觸前膜內的負電位,便於小泡與突觸前膜接觸而發生融合。小泡破裂把遞質和其他內容物釋放到突觸間隙時,其外殼仍可留在突觸前膜內(也可與突觸前膜融合,成為突觸前膜的組成部分),以後仍舊可以重新恢復原樣,繼續合成並貯存遞質。
從突觸小泡的胞吐作用到小泡膜的回復可分為下列6個時相:
①突觸小泡靠進突觸前膜活性帶;
②小泡貼靠突觸柵欄結構;
③小泡與突觸前膜接觸和兩膜融合;
④融合膜裂開向突觸間隙釋放神經遞質;
⑤小泡膜併入突觸前膜質;
⑥小泡膜回收並重新利用。在小泡膜的循環過程中,有一些膜不形成功能性小泡不進入循環而是被溶酶體降解並通過逆向軸漿運輸返回胞體重新加工。同時通過順向軸漿運輸將新的小泡送往神經終末。
神經遞質由突觸前膜釋放后立即與相應的突觸后膜受體結合,產生突觸去極化電位或超極化電位。導致突觸后神經興奮性升高或降低。自此,神經衝動的電信號就完成了對突觸間的一次跨越。
突觸處遞質釋放過程
進入突觸間隙的乙醯膽鹼作用於突觸后膜發揮生理作用后,就被膽鹼酯酶水解成膽鹼和乙酸,這樣乙醯膽鹼就被破壞而推動了作用,這一過程稱為失活。去甲腎上腺素進入突觸間隙併發揮生理作用后,一部分被血液循環帶走,再在肝中被破壞失活;另一部分在效應細胞內由兒茶酚胺內由兒茶酚胺位甲基移位酶和單胺氧化酶的作用而被破壞失活;但大部分是由突觸前膜將去甲腎上腺素再攝取,回收到突觸前膜處的軸漿內並重新加以利用。多巴胺的失活與去甲腎上腺素的失活相似,它也是由兒茶酚胺氧位甲基移位酶和單胺氧化酶的作用而被破壞失活。突觸前膜也能再攝取多巴胺加以重新利用。5-羥色胺的失活也與去甲腎上腺素的失活相似,單胺氧化酶等能使5-羥色胺降解破壞,突觸前膜也能再攝取5-羥色胺加以重新利用。氨基酸遞質在發揮作用后,能被神經元和神經膠質再攝取而失活。肽類遞質的失活是依靠酶促降解,例如通過氨基肽酶、羧基肽酶和一些內肽酶的降解而失活。
基本信息
- 中文名
- 神經遞質
- 別名
- 遞質
- 外文名
- neurotransmitter
- 作用
- 信使
- 屬於
- 生物學
- 傳遞
- 突觸傳遞
- 分泌方式
- 旁分泌
- 釋放位點
- 突觸前膜
- 作用位點
- 突觸后膜