定向
生物行為
定向是動物主動調整其身體或身體某部分的空間位置的行為。
動物主動調整其身體或身體某部分的空間位置的行為。如青蛙伸舌粘取飛蟲的取食行為,以及候鳥往返數千里飛翔于越冬及繁殖地的遷徙,就其需要精確地確定方向而言,都屬於典型的定向行為。定向行為的對象可為外物,如鳥獸類使所築巢穴有一定朝向(如避風朝陽)。空間位置可以指靜止的位置,如動物休息時哪一部分貼附於基底都有定規。又如在陽光下,蜥蜴常根據外界溫度而採取一定的體位方向使所吸收的日光足以保持體溫大致穩定。再如猛獸隱藏樹叢中準備伏擊過路動物,必須維持朝向一定方向的警覺待動的姿態。但動物還要不斷地活動,要覓食、求偶、避敵等等,這都存在定向問題。動物的捕食動作不僅要敏捷有力,而且一定要對準目標。飛鳥四齣尋食路途迂迴,甚至會遇到風雨,但卻能順利返巢。這一切的關鍵在於動物能辨別方向。在定向行為的全過程中隨時都要辨別方向,校正可能發生的偏差。
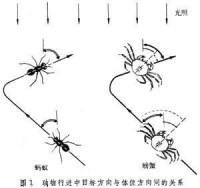
動物的靜止姿態和活動路線大都有明確取向。動物的四齣覓食似無目的,但實際上有內在規律,或循氣味、聲響等線索,或採取某種程序按區搜索。例如螞蟻在植葉上常是沿某一邊線(或左或右)循序查遍各區。再如海邊的招潮蟹的巡視路線很象一朵花,自同一點出發沿輻射方向外出,畫一個小弧回來,再沿另一輻射向外出,如此循序繞遍幾個方向。方向是就一定的參考系統而言。可能是針對一個點,例如捕食的目標、求偶的對象、返歸的巢穴等;也有的是就某個方面而言,例如沿重力線有上下之分,上下本來只是相對而言,但若劃定一個界面(如地面或水面),便有了絕對的分界。不過在一般描述中總採用一定的坐標系統。在左右對稱的動物中,可以利用3組剖面將身體分為頭尾、腹背和左右幾個方面。在橫剖面上,以頭尾軸作中心還可分內外方向。至於動物在環境中的位置,則一般總是採取地理坐標,即東西南北方位。體位的定向 動物不活動時的體位大多是相對於重力線、基底或外界光線而保持一定方向。在傾斜的地面上,體重較大的動物總力圖保持其垂直軸與重力線一致。但重力對微小動物的影響就不如基底的影響大。例如家蠅在頂板上和地面上的行動同樣自如。許多水生動物因水浮力的關係,受重力的影響較小,而頂側來的天光成為其重要的定向參考因素,這些動物總是保持背向天光的體位。再因為它們背側皮色較深而腹側較淺,所以與頂側天光配合,還有利於隱形避敵。大多數動物行動時,身體頭尾軸與路線方向一致;但螃蟹行動時,不管路線如何迂迴,其身體卻保持與環境成一固定角度,因此身體方向與路線方向並不一致。
對於短距離行動,地形或氣味標記,便可引導動物直達目的地;而遠距離航行則需要其他線索導向。從理論上講,地球上兩點相距不論多遠,要在其間直線航行只需要知道方向和距離。只要取向正確,動物航行中所消耗的體力可有助於估計走過的距離,而局部地形則有助於辨識是否已到達目的地。因此定向就成為關鍵問題。僅僅知道東西南北各在哪方不行,要知道的是目的地在自己的哪一方。對於人類飛行來講,只需要知道起飛地和目的地各自的經度緯度,便可在地圖上找出方向和距離。但動物無相應機制。目前所能肯定的只是很多動物能以太陽或其他天體為定向的指針,不過這也只是指出動物四周的地理方位,並不能據此判定目的地在何方。對於地表觀察者來說,天體位置是隨時間而有規律地變動著,因此,還要藉助定時能力來判斷方位。
動物的行為都是內外因子的綜合結果,內部生理狀態產生行為的動因,外在環境因子則觸發具體行動。對於定向行為來說,還存在導向因子。例如小鼠見貓急奔回穴,貓是奔跑行為的觸發因子,洞穴則是導向因子。而貓捕鼠時,鼠對於貓來說,既是觸發因子,又是導向因子。
定向
來自外界導向因子的刺激,也可大致分為3類:機械的、化學的和輻射的。機械性刺激包括觸壓覺、平衡覺和聽覺等。龍蝦的螯肢上覆有一層感覺性纖毛,纖毛的不同排列可以探測不同方向的水流,這些纖毛組成一個屏幕式感覺器官,足以辨別水流方向的種種變化。平衡覺一般是辨別重力方向以及各種加速運動的方向,幫助調整身體姿勢。機械刺激中,以波動最有助於定向。仰蝽可根據水波方向發現附近的獵物,這屬於低於聲頻的次聲波。高於聲頻的超聲波,方向性更強,藉此可在黑暗中辨物。蝙蝠能在深夜或無光線的岩洞中捕捉飛蟲,就是靠本身發出超聲束再借感官接受反射回的超聲波辨向的。其原理近於雷達,被稱為聲納。
所謂化學性刺激主要是通過嗅覺感知的各種氣味。食物、配偶或天敵都可散發出特殊氣味,形成一個自內向外由高到低的濃度場。動物可借濃度梯度辨彆氣味源的方向。例如,螞蟻常在取食路途上散布特定的分泌物,供同巢螞蟻循跡跟蹤。另外,氣味隨氣流運動,故逆風而行是很多動物辨向覓食的重要方式。
動物可以感覺遠距離發來的輻射,而且定位準確。例如,在人類的視覺範圍之外,響尾蛇可察覺微弱的紅外輻射;某些昆蟲可感受紫外輻射以及光線的偏振方向;有少數魚類可以在水中產生微弱電場,藉此察覺一切游入電場內的其他生物;一些鳥類可以感覺天然磁場,歐歌鴝在天然磁場下採取一定方向,但在附加人工磁場后,即轉而採取兩者的合力方向。
一切遠出覓食后返巢飼雛的鳥獸都必須具有一定的辨向能力。蜜蜂離巢數百米覓食時,盤繞迂迴飛行,而返巢時卻可直線飛回,說明它能判斷巢在自己的哪一方。也有令人難理解的例子,如鮭、鰻等遠距離遷游和候鳥遠距離遷飛是如何定向的。人們研究最多的是信鴿,將信鴿運至近千公里之外再釋放,不到一天就可返回家。在陌生地點,它開始時只是繞圈飛,但越繞越大,然後便以近於直線的路程返家,直到發現熟悉一些的地形標記時才最後調準路線直抵家中。有些地面動物行走時沿途以分泌物標記,故可以循原路返回。實驗證明,細腰蜂可以辨識巢穴周圍的地形標記。可能很多動物在日常活動中是依靠辨識地形定向的,如信鴿在接近目的地時是如此。很多候鳥常沿河流、山脈、海岸線遷徙,可能便是把地形作為導航的一個手段。但無論氣味標記或地形辨識都只能引導動物重走舊路,而不能解釋信鴿在陌生地點被釋放時的定向行為。
動物在遠航時可能同時採用多種導航手段,在航線的不同階段重點有所不同。各物種之間採用的手段也不相同,這與它們所具有的感覺能力有關。遠航前,動物需要知道的不僅僅是東南西北,而是目的地的確切方向,在航行中還要能保持這個方向,不論是氣流、水流或地形障礙使它偏移航向多遠,它都必須能及時發覺和糾正。在這方面,地形及視覺辨識只能起輔助作用。有人曾提出,地心引力和因地球自轉造成的科里奧利氏力都隨緯度而變,動物可能藉此判斷自身所處緯度。但實驗未能證實這個設想。某些動物確能感覺地球磁場,可能以此作為導航手段,但這也只能決定南北方向的位置。
定向
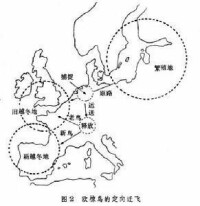
歐洲園鶯自幼在籠中飼養,至秋季遷飛時節經常面向西南方(習慣遷飛的方向)。這說明定向行為有其遺傳基礎。但波羅的海一帶繁育的椋鳥的傳統越冬地在英格蘭和愛爾蘭的南部及法國北部。在秋季,當它們途經荷蘭時加以截獲並套環標記,再送至瑞士釋放。這樣通過連續幾年的實驗。結果發現初次飛行的新鳥仍按習慣的地理方向飛行竟飛到西班牙一帶去越冬,而飛過一次以上的老鳥卻能改變航向,仍飛到傳統越冬地(圖2)。這表明,學習與經驗有助於確定目標方向。
動物的各種行為都有針對的目標,因而都有定向問題。無脊椎動物中,定向能力以昆蟲最為發達。脊椎動物中,則以魚類、鳥類和哺乳類中一些年年遠距遷徙的物種表現得最為突出。而且隨著環境介質不同,動物演化出不同的定向手段。陸地上,氣味標記是較古老的方法。在黑夜,在地穴和海洋中,某些飛行的和水生的哺乳動物演化出超聲定向的本領。但定向能力最發達的還要屬鳥類。有的候鳥能飛翔萬里,具有敏銳的視力足以辨識大面積的地形特徵。很多候鳥還具有生物鐘,因此能借太陽或星辰指向,甚至在濃雲密布的天氣中,它還可借磁場來保持航向。

基本信息
- 類別
- 生物行為
- 原因
- 內外因子的綜合結果