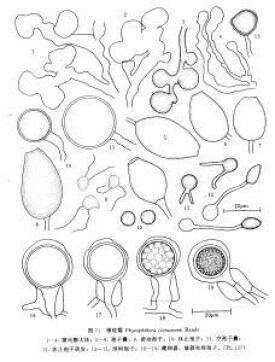
樟疫霉
樟疫霉
樟疫霉在CA培養基上菌落綿絮狀,氣生菌絲中等到繁茂,邊緣不明顯。
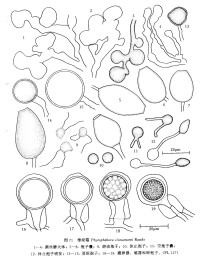
樟疫霉
鱷梨Persea americana Mill.:雲南西雙版納(PL 137)。
刺槐Robinia pseudoacacia L. :江蘇贛榆 [HP31(A1),南京(P72)]。
黃世珏和王建國(1983)沈崇堯等(1989)和徐敬友等(1990)分別報道此菌在江蘇和北京為害刺槐。何漢興等(Ho et al., 1984a)、陸家雲等(1984)、陸家雲和鄭小波(1988)以及徐敬友等(1989,1990a)報道在江蘇、浙江、福建為害雪松Cedrus deodara (Roxb.)D. Don、鱷梨和山茶Camellia japonica L. 等。吳玲忠(1989)報道在上海地區為害茶屬Camellia sp.、鳳仙花Impatiens balsamina L.、杜鵑花Rhododendron hy- bridum Hort.、香樟Cinnamomum camphora Presl和木瓜 Chaenomeles sinensis (Thouin) Koehne等。沈崇堯等(1989)首次在我國發現P. cinnamomi A2於茶花上。
在我國台灣,澤田(Sawada,1942)報道此菌寄生於李氏金雞納樹Cinchon ledgerina Moens和金雞納樹C. succirubra Pav.。張和喜(1977)、何漢興(Ho,1990)和蔡雲鵬(1991)報道此菌在台灣的寄主還有:鳳梨Ananas cosmosus(L.)Merr. 、山茶Camellia japonica L. 、柑橘類、中美木棉Pachira macrocarpa( CHAM. & Schlecht.)Schlecht. ex Bail.、鱷梨、杜鵑花屬Rhododendron sp. 等。
幾內亞、土耳其、馬爾加什、馬來西亞、中國、日本、扎伊爾、烏干達、烏拉圭、牙買加、比利時、巴貝多、巴西、巴布亞紐幾內亞、巴拿馬、以色列、加拿大、印度、印度尼西亞、聖克羅什、聖偉省提、古巴、西班牙、剛果、多米尼加、瓜地馬拉、庫克島、貝里斯、希臘、英國、法國、肯亞、羅得西亞、波多黎各、阿根廷、委內瑞拉、南非、南斯拉夫、喬治亞、玻利維亞、宏都拉斯、海地、美國、哥倫比亞、哥斯大黎加、愛爾蘭、荷蘭、秘魯、特立尼達、斐濟、菲律賓、薩爾瓦多、俄羅斯、葡萄牙、越南、象牙海岸、智利、喀麥隆、瑞士、義大利、紐西蘭、摩洛哥、澳大利亞、墨西哥、德國、尚比亞。
Phytophthora cinnamomi Rands分佈甚廣,全世界約有70個國家、地區和島嶼均有報道,它侵染273屬900多種不同的寄主植物,A2交配型比A1交配型分佈為廣,A1僅分佈在中國、澳大利亞、美國、南非、馬爾加什和巴布亞紐幾內亞等地(Ribeiro,1978;Zentmyer, 1980;Ho et al.,1983,1984;陸家雲、鄭小波, 1989;徐敬友等,1989,1990a)。本種不同交配型的形態也有差異:A1交配型的孢子囊小,多為檸檬形;A2 交配型孢子囊較大,多為倒梨形(Shepherd et al.,1974;Ho & Zentmyer, 1977)。在美國夏威夷兩種交配型均有(Ko et al.,1978;Ko,1979)。在我國東南沿海和台灣等省,A1 和A2兩種交配型均有發現,江蘇、上海、浙江、福建以A1交配型佔優勢。
Phytophthora cinnamomi Rands並不是原產於美國加州或拉丁美洲的土壤里(Zent- myer,1976,1977),據信也不是原產於澳大利亞和紐西蘭(Newhook & Podger,1972),它是在18世紀引入這些地區的。但是,Shepherd等( 1974)認為此菌是在一百多方年以前從新澳大利亞傳入新幾內亞、西里伯斯(Celebes)地區的。 Zentmyer(1976,1977)還認為P. cinnamomi是從馬來西亞凍澳大利亞地區在早年的航海和探險中,通過太平洋和美洲,伴隨植物材料和土壤首先傳入西半球的。他相信此菌是在19世紀末 20世紀初,由夏威夷和中美洲的鱷梨和其他熱帶植物的引種而傳到美國加州的。柯文雄等(Ko et al.,1978)研究了台灣此菌的交配型 A1:A2=42:30,接近1 :1,因此他們認為此菌起源於亞洲,中國台灣被包括在起源中心之內。
Royle & Hickman(1964)和Ho & Zentmyer(1977)都曾觀察到有些Phytophthora cinnamomi Rands的雄器是雙細胞的,但我們研究的菌系(如PL 137)的雄器卻為單細胞。
Hwang et al.(1978)報道Phytophthora cinnamomi Rands在土壤中的壽命:厚垣孢子存活最久,孢子囊次之,遊動孢子最為短暫。