木霉
木霉
木霉屬於半知菌門,絲孢目,木霉屬,常見的木霉有綠色木霉、康寧木霉、棘孢木霉、深綠木霉、哈茨木霉、長枝木霉等。
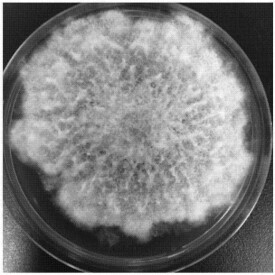
木黴菌落開始時為白色,緻密,圓形,向四周擴展,后從菌落中央產生綠色孢子,中央變成綠色。菌落周圍有白色菌絲的生長帶。最後整個菌落全部變成綠色。綠色木黴菌絲白色,纖細,寬度為1.5~2.4微米。產生分生孢子。分生孢子梗垂直對稱分歧,分生孢子單生或簇生,圓形,綠色。綠色木黴菌落外觀深綠或藍綠色;康氏木黴菌落外觀淺綠、黃綠或綠色。
【拉丁學名】Trichoderma spp.
綠色木霉分生孢子梗有隔膜,垂直對生分枝;產孢瓶體端部尖削,微彎,尖端生分生孢子團,含孢子4~12個;分生孢子無色,球形至卵形,2.5~4.5×2~4μm。
綠色木霉適應性很強,孢子在PDA培養基平板上24℃時萌發,菌落迅速擴展。培養2天,菌落直徑為3.5~5.0厘米;培養3天,菌落直徑為7.3~8.0厘米;培養4天,菌落直徑為8.1~9.0厘米。
深綠木黴菌塗片標本採集時間及情況:採集點為木板改料露天現場廢料與新木製樹盆表,採集時間:20160429 8:25AM室內溫度23℃,室外溫度23-(3至5)℃,夜雨後,濕度53%。孢子浸潤松木板材木表:菌落肉眼觀呈深綠色,藍綠色,色斑直徑多在1-2mm,大者條狀4*13mm,更甚冠狀面觀測孢子浸潤整塊松木板材的近樹軸側,年輪越小越容易導致綠色木霉整片浸潤,可能與木材冠狀切面的年輪起到了菌絲生長導向和限制作用有關。採集4小時后25*100*2顯微油鏡觀察發現:孢子1-2mm鏡下物呈散囊靜止狀態,深綠色或藍綠色,金屬光澤,孢子居多,菌絲白色或深綠色少,菌絲分節。
通常菌落擴展很快,特別在高溫高濕條件下幾天內木黴菌落可遍布整個料面。菌絲生長溫度4—42℃,25—30℃生長最快,孢子萌發溫度10—35℃,15—30℃萌發率最高,25—27℃菌落由白變綠只需4—5晝夜,高溫對菌絲生長和萌發有利。孢子萌發要求相對濕度95%以上,但在乾燥環境也能生長,菌絲生長pH值為3.5~5.8,在pH值4~5條件下生長最快。
危害癥狀木霉的主要生物特徵為其菌絲成熟期很短,往往在一周內即可達到生理成熟,然後即生出綠色霉層,即其孢子層。當基料被侵染后,菌絲階段不易察覺,直到出現霉層時才能引起注意;起初只是點狀或斑塊狀,當條件合適或食用菌菌絲不很健壯時,很快發展為片狀,直至污染整個菌袋或料床,若不及時採取措施,菇棚內短時間即可成一片綠色,其孢子飛揚,周邊棚牆上也將附著大量木霉孢子,給以後的生產留下嚴重隱患。
發生規律木霉主要生存在朽木、枯枝落葉、土壤、有機肥、植物殘體上和空氣中。許多栽培的老菇房,帶菌的菇具和場所是主要的初侵染源,已發病所產生的分生孢子,可多次重複侵染更為頻繁。木霉發病率的高低與環境條件的關係較大,木霉孢子在15-30`C下萌發率較高,菌絲體在4-42`C的溫度下均能生長,在25-30`C生長最快。孢子在空氣相對濕度95%的條件下,萌發最快,相對濕度低於85%較難萌發。因此,在高溫、高濕、通氣不良和培養料呈偏酸性時,很容易滋生木霉。木霉侵染寄主后,與寄主爭奪養分和空間,同時還分泌毒素殺傷、殺死寄主,把寄主的菌絲纏繞、切斷。
(1)制種或熟料栽培拌料時按比例加入1:1000倍疣霉凈,並嚴格滅菌,以徹底殺死其孢子。
(2)科學調配基料組分,使營養全面、均衡,以保證食用菌菌絲的健康和抗性,可對黴菌形成拮抗或抑制。實踐證明,生產中按比例加入天天菇耳壯即可。
(3)發酵栽培時,加入疣霉凈后,基料仍要發酵均勻,儘可能多的殺死或抑制其孢子。
(5)菌種或菌袋發菌以及出菇期間,每5天左右對菇棚空閑處噴灑30-50倍金星消毒液或1:1000倍疣霉凈,是目前最有效的預防方法。
(6)發現木霉后,及時用1:500倍疣霉凈噴灑或注射、塗抹污染區和菌袋,效果顯著,污染嚴重的菌袋要及時作焚燒或深埋處理。
木黴菌可用來防治病害或抑制病原的主要機制,其行為通常可歸類成五大類,即產生抗生素、營養競爭、微寄生、細胞壁分解酵素、以及誘導植物產生抗性。一般而言,上述機制雖會因木黴菌種類或菌株的不同而出現主要功能上的差異,但病害防治的整體機制通常會涵蓋一種以上。
產生抗生素
木黴菌可以產生揮發性或非揮發性抑制病原菌生長的抗生物質,如三柯勝、三柯得茗、粘帚毒素、煤尼毒素及胜肽素等。司齊博克(Mary Schirmbock)等人曾觀察到哈氏木黴菌可產生細胞壁分解酵素及胜肽博素的抗生素,如果把這種抗生素與幾丁質分解酵素結合,可抑制病原菌孢子發芽與菌絲生長。
營養競爭
利用競爭能力強的微生物,消耗如鐵、氮、碳、氧或其它適宜病原菌生長的微量元素,可以限制病原菌的生長、發芽或代謝。在這方面,木黴菌主要是奪取或阻斷病原菌所需的養分。由於營養競爭很難用變異菌株加以證明,而且添加物質也可能會改變病害的發生,以致無法取得強而有力的證據,顯示防治的機制是與競爭養分有關。目前較具明證者,僅在鐵、銅等離子的競爭方面,而這又與能否產生嵌合物質等具有相關性,因為這類物質也會減少病原菌的發芽與生長。
細胞壁分解酵素
一般認為細胞壁分解酵素在抑制病害上扮演著重要的角色。由於幾丁質與葡聚糖是真菌細胞壁的主要成分(除卵菌綱外),很多試驗顯示幾丁質分解酵素或葡聚糖分解酵素,單獨或組合使用時可直接分解真菌細胞壁。近來遺傳學上也證明,缺乏幾丁質分解酵素的突變菌株,其抑制病原菌孢子發芽的能力以及病害防治能力都明顯降低。
試驗顯示,如果把幾丁質分解酵素基因引入無病害防治能力的大腸桿菌菌株中,這個轉殖菌株就可減少大豆白絹病的發生。同樣地,把來自薛利蒂亞細菌的幾丁質分解酵素引入到哈式木黴菌菌株后,這種菌株也比原來菌株具有更強的覆蓋白絹病生長的能力。最近更有很多轉殖植物含有來自木黴菌的幾丁質分解酵素,因而增加了它們對植物病原真菌的抗性。
微寄生
以木黴菌的微寄生立枯絲核病菌為例,其過程大約可分成四個步驟。首先是趨化性生長,也就是木黴菌會趨向能產生化學刺激物的病原菌生長。第二個步驟是辨識,這個步驟和病原菌含有的聚血素及拮抗菌表面擁有的碳水化合物接收器有關,這類物質左右了病原菌與拮抗微生物之間作用的專一性。第三個步驟是接觸與細胞壁分解。最後則是穿刺作用,也就是木黴菌會產生類似附著器的構造,侵入真菌細胞,進而分解與利用病原菌細胞物質。
誘導植物產生抗性
植物的系統性誘導抗病現象,是指植物經第一次接種原或非生物因子刺激后,產生對第二次接種原的抗性。這種抗性的發展,可導致植株對多種病原的感染都會有抵抗性,而非僅限於對原先的誘導病原。目前已有報告顯示,植物經木黴菌處理后,可誘導產生特別的酵素等物質,進而對葉部病害或病毒病害產生抗性。
基本信息
- 中文名
- 木霉
- 外文名
- Trichoderma spp.
- 界
- 真菌界
- 亞門
- 盤菌亞門
- 門
- 半知菌門
- 綱
- 糞殼菌綱
- 亞綱
- 肉座菌亞綱
- 目
- 絲孢目
- 科
- 肉座菌科
- 屬
- 木霉屬
- 體征
- 木黴菌落開始時為白色,緻密,圓形,向四周擴展,后從菌落中央產生綠色孢子,中央變成綠色