多基因
多基因
對於同一性狀的表型來講幾個非等位基因中的每一個都只有部分的影響,這樣的幾個基因稱為累加基因或多基因。
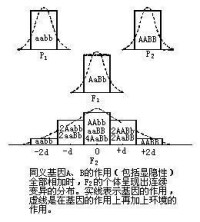
多基因
1908年尼爾遜·埃爾提出多基因假說對數量性狀的遺傳進行了解釋。按照他的解釋,數量性狀是許多彼此獨立的基因作用的結果,每個基因對性狀表現的效果較微,但其遺傳方式仍然服從孟德爾的遺傳規律。而且還假定:
(1)各基因的效應相等。
(2)各個等位基因的表現為不完全顯性或無顯性,或表現為增效和減效作用。(3)各基因的作用是累加的。
我國人口眾多、遺傳資源豐富,多基因病研究具有獨特的優勢。在對中國群體遺傳結構研究的基礎上開展我國多基因病相關基因的研究課題日益得到重視。本課題對來自中國各地的9988例男性隨機樣本進行了M89、M130和YAP三個Y染色體單倍型的基因分型。這些位於Y染色體非重組區的突變型M89T、M130T和YA屍均來自另一Y染色體單倍型M168T,M168是除非洲以外所有現代人及部分非洲人共同具有的一種古老的突變型,在除非洲以外地區沒有發現一例個體具有比M168更古老的突變型。結果顯示近萬份樣品無一例外具有這三種突變型之一,這一結果為現代人群的非洲起源假說提供了重要佐證,並認為Y染色體的證據並不支持即使是對現代中國人起源可能起著極小作用的獨立起源假說。同時通過全基因組掃描對鼻咽癌高癌家系和原發性低血壓家系進行家系連鎖分析,鼻咽癌研究結果初步顯示在2號、6號和Q號等染色體存在支持連鎖的區域,其中 D6S422位點Lodscore(Z2all)=l 198874、NPL=2.33113(P‘0 00684),原發性低血壓家系研究中發現2、7和11號染色體存在支持連鎖區域,其中D2S112,Lod score(Z a11)=1.12986,NPL=2.25152(p=0.00799),D11S901位點Lod score(Z a11)=2.089601、NPL= 2.37206(p=0.00576)均表現顯著性的信號。部分結果得到過去一些研究成果的訐實,這些數據為進一步通過候選定位克隆研究進行鼻咽癌和血壓調控相關基因提供了重要實驗依據。