胞間連絲
胞間連絲
在初生紋孔場上集中分佈著許多小孔,細胞的原生質細絲通過這些小孔,與相鄰細胞的原生質體相連。這種穿過細胞壁,溝通相鄰細胞的原生質細絲稱為胞間連絲。是細胞間物質運輸與信息傳遞的重要通道,通道中有一連接兩細胞內質網的連絲微管。
胞間連絲(plasmodesmata)
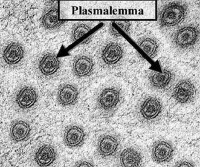
電鏡下胞間連絲的正面觀

胞間連絲
胞間連絲(plasmodesma)貫穿兩個相鄰的植物細胞的細胞壁,並連接兩個原生質體的胞質絲。它們使相鄰細胞的原生質連通,是植物物質運輸、信息傳導的特有結構這一結構由E.坦格爾於1879年首先在馬錢子胚乳細胞間發現。1882年由德國植物學家、細胞學家E.A.施特拉斯布格命名為“胞間連絲”。近年來的一個重要的進展就是,發現胞間連絲運輸的物質中,不僅僅是一些諸如礦質離子、糖、氨基酸和有機酸等小分子物質,而且含有諸如蛋白質和核酸等大分子物質。日益增多的大量證據表明,胞間連絲通道對運輸物質分子大小的限度也受許多因子的調節。
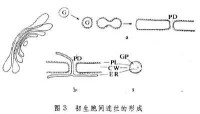
胞間連絲
胞間連絲與動物細胞的間隙連接有許多相同之處。正常情況下它允許1000道爾頓以下的分子滲透,也能讓離子自由通過。它的活性同樣受Ca2+離子濃度的調節等,因此具有植物信號傳導的作用。與間隙連接不同的是,胞間連絲的孔能夠擴張,允許大分子,包括蛋白質和RNA分子通過。
多樣性
胞間連絲多見於高等植物,某些藻類以及真菌亦有存在,較多的出現在紋孔的位置上。植物體的各個細胞正是通過胞間連絲,彼此相互形成統一整體的。但某些成熟細胞之間有時並不存在這種結構。在同一細胞的不同部分的壁上,胞間連絲出現的數目常有不同。
現今一種包含壓扁內質網(ER)的胞間連絲的模式已被確認,這種結構模式較過去有一個新的發展,胞間連絲通道周圍是相鄰兩個細胞的質膜的延續和連接,其中央有一個由壓扁ER形成的圓筒體,名為連絲橋管。一種約3nm的蛋白質顆粒包埋在其周圍質膜和中央橋管-ER膜中,另一種電子稠密的輻射狀纖絲連接著這兩者中的蛋白質顆粒。包埋在中央橋管ER膜上的蛋白質顆粒呈現螺旋式或一系列圓圈式旋轉排列,在橫切面上,可以看見7-9個顆粒。連接中央橋管外側和質膜內側兩者膜上的蛋白顆粒的輻射狀纖絲可能是肌動/肌球蛋白和激酶。這種纖絲的長度約為2.5nm,這也就是胞間連絲通道運輸的量度和限度。
近些年,在對冬小麥幼葉組織胞間連絲的研究中,發現至少有4種類型的胞間連絲存在於這種幼葉組織的細胞壁中。這四種類型是:一種是典型的包含壓扁的ER,並顯示明顯的“頸”結構的胞間連絲,在這種胞間連絲的中部腹區中央可清晰地看到壓扁的ER。另一種是,一種直行通道的胞間連絲,沒有明顯的“頸區”,但也包含壓扁的ER,然而這種中央橋管與周圍質膜的聯繫似乎比較鬆散,其通道的運輸量度可能較大。第三種是,分枝形的胞間連絲,其中央也含有壓扁的ER。第四種是,一種僅為相鄰細胞間連續質膜包含的通道,其中沒有壓扁的ER,這種簡單的胞間連絲通道一般比較大。這一結果進一步的節食和證明,那種包含壓扁ER、具有“頸”型的胞間連絲不是高等植物胞間連絲的惟一結構模式。
胞間連絲的主要功能是:①細胞間物質包括小泡的運輸和轉移;②信息、刺激的傳導;③影響細胞的生長、發育和分化。如高等植物頂端分生組織的胞間連絲分佈狀況就與分化的控制有關,例如蕨類植物桂皮紫萁胞間連絲在垂周壁與平周壁的分佈數目不同就影響了頂端早期葉的發育。此外病毒有時也可經胞間連絲傳播。
胞間連絲
次生胞間連絲是由一些降解酶(果膠酶、半纖維素酶和纖維素酶)的作用使完整的細胞壁穿孔而成。被子植物花粉母細胞間減數分裂早前期出現的次生胞間連絲直徑大,約0.5~1.5微米,特稱細胞融合道。雖然它們能在初生的胞間連絲的位置上形成,但和一般胞間連絲並不完全相同。
胞間連絲通道的調節
通過熒光染料擴散實驗指出,胞間連絲容許通過物質分子大小限度(SEL)一般為800-1000Da。近年來的研究結果揭示,這種SEL與器官組織細胞的功能有關。Nicotiana clevelandii表皮毛細胞間的胞間連絲SEL可達7000Da;Cucurbita maxima和Vicia faba莖的篩管分子和伴胞之間的胞間連絲SEL分別為3000和10000Da;並揭示胞間連絲通道口徑的開放程度受許多因子的調節。
大量實驗結果指出,Ca2+是胞間連絲口徑的重要調節者。通過顯微技術向細胞內注入Ca2+偶聯熒光染料,結果顯示,在細胞含有高濃度Ca2+的條件下抑制了熒光染料進過胞間連絲在細胞間的擴散;而在對照細胞中(只注入熒光染料,沒有結合Ca2+),則熒光染料可以通過胞間連絲從一個細胞擴散到另一個細胞。巨型輪藻在冬季里,由於細胞內有較高的Ca2+水平,細胞間交通受阻,生長停止,處於休眠;到春季,細胞內的Ca2+濃度降低,胞間通道通暢,生長恢復。用含有Ca2+載體的溶液培養,或直接注入Ca2+溶液,提高春天輪藻細胞的Ca2+濃度,其胞間交通又返回冬季時機的狀態。細胞內的ATP的含量也起著調節連絲口徑的作用。Cleland等通過疊氧化物和缺氧脅迫,降低細胞內ATP含量,結果使小麥根細胞連絲通道的排阻分子限度(SEL)從小於800Da增加到7-10kDa。這進一步揭示了胞間連絲通道受控於ATP依賴的磷酸化。
1,胞間連絲把構成植物體基本結構單位的細胞形成為一種辯證統一體:它使這些多細胞彼此溝通,相互連接;但又保持各細胞的相對獨立,仍然是生命活動的一個基本單位,具有遺傳上的全能性。
2,通過胞間連絲形成的共質體不是一成不變的,為適應植物體生長與發育發生不斷的改變和重新構建。
3,已知的胞間連絲調節因子有;類括約肌,肌動/肌球蛋白胼坻質及Ca2+等。
基本信息
- 中文名
- 胞間連絲
- 外文名
- plasmodesmata
- 類型
- 醫學名詞
- 特點
- 具有多樣性
- 功能
- 某些細胞進行細胞間交流的載體
- 常見地點
- 高等植物、某些低等植物