共找到2條詞條名為生態因子的結果 展開
- 生態因子
- 生態因素
生態因子
生態因子
生態因子(ecological factor)指對生物有影響的各種環境因子。常直接作用於個體和群體,主要影響個體生存和繁殖、種群分佈和數量、群落結構和功能等。各個生態因子不僅本身起作用,而且相互發生作用,既受周圍其它因子的影響,反過來又影響其它因子。
植物的環境是一個廣義的概念。對於具體的植物和植物群落,我們一般使用“生境”這一概念。
植物的生境包括許多環境要素,如大氣濃度一項就包括氧氣的濃度、二氧化碳的濃度、惰性氣體的濃度等。但並不是所有的環境要素都對植物的生活產生影響,如惰性氣體的濃度基本上不影響植物的生活。我們把對植物有影響的,直接作用於植物生命過程的那些環境要素稱為生態因子,又稱生態因素。
生態因子分為非生物因子、生物因子和人為因子三大類。非生物因子主要包括氣候因子(如光照、溫度等)、水分因子和土壤因子等。生物因子主要指植物之間的機械作用,共生,寄生,附生,動物對植物的攝食、傳粉和踐踏等。人為因子包括人類的墾殖、放牧和採伐,環境污染等,是一類非常特殊的因子。
在各種生態因子中,並非所有的因子都為植物的生長所必需。我們把植物生長所必需的因子稱為生存條件,亦即植物缺少它們就不能生長。對於綠色植物來說,這些因子是氧氣、二氧化碳、光、熱、水和無機鹽。
當環境中某種生存條件出現異常,便會抑制植物生命活動或威脅植物生存,這種現象稱為環境脅迫。動物嚙食、寄生、風害、火災和土壤侵蝕等現象可以部分或全部地破壞植物生命活動的產物,被稱為干擾。
生態因子
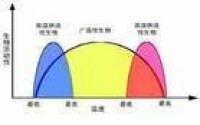
後來最低量律被擴大到包括植物和動物的各種環境要求。另一方面,某種生態條件(物質或能量)太多也同樣起限制作用。一般說來,生物對於大多數生態因子有一定的耐受極限耐受上限和耐受下限。1913 年美國動物學家 V.E.謝爾福德曾把這一概念稱作耐受原理,即某類生物的多度或分佈被超過該生物所能耐受的限和最低限的因子所控制。
生態因子的類型多種多樣,分類方法也不統一。簡單、傳統的方法是把生態因子分為生物因子(biotic factor)和非生物因子(abiotic factor)。前者包括生物種內和種間的相互關係;後者則包括氣候、土壤、地形等。根據生態因子的性質,可分為以下五類:
氣候因子也稱地理因子,包括光、溫度、水分、空氣等。根據各因子的特點和性質,還可再細分為若干因子。如光因子可分為光強、光質和光周期等,溫度因子可分為平均溫度、積溫、節律性變溫和非節律性變溫等。
土壤是氣候因子和生物因子共同作用的產物,土壤因子包括土壤結構、土壤的理化性質、土壤肥力和土壤生物等。
地形因子如地面的起伏、坡度、坡向、陰坡和陽坡等,通過影響氣候和土壤,間接地影響植物的生長和分佈。
生物因子包括生物之間的各種相互關係,如捕食、寄生、競爭和互惠共生等。
把人為因子從生物因子中分離出來是為了強調人的作用的特殊性和重要性。人類活動對自然界的影響越來越大和越來越帶有全球性,分佈在地球各地的生物都直接或間接受到人類活動的巨大影響。
生態因子的劃分是人為的,其目的只是為了研究或敘述的方便。實際上,在環境中,各種生態因子的作用並不是單獨的,而是相互聯繫並共同對生物產生影響,因此,在進行生態因子分析時,不能只片面地注意到某一生態因子,而忽略其他因子。另一方面,各種生態因子也存在著相互補償或增強作用的相互影響。生態因子在影響生物的生存和生活的同時,生物體也在改變生態因子的狀況。
美國植物生態學家道本邁爾(R.F.Daubenmire)將生態因子分為七個並列的項目:土壤、水分、溫度、光照、大氣、火和生物因子。在研究園林植物與環境的相互關係時,主要考慮除火因子之外的其他六個,因為它們常常直接影響園林植物的生長發育。

生態因子
綜合性
生態因子
非等價性
對生物起作用的諸多因子是非等價的,其中有1~2個是起主要作用的主導因子。主導因子的改變常會引起其他生態因子發生明顯變化或使生物的生長發育發生明顯變化,如光周期現象中的日照時間和植物春化階段的低溫因子就是主導因子。
不可替代性和可調劑性
生態因子雖非等價,但都不可缺少,一個因子的缺失不能由另一個因子來代替。但某一因子的數量不足,有時可以由其他因子來補償。例如光照不足所引起的光合作用的下降可由CO2濃度的增加得到補償。
階段性和限制性
生物在生長發育的不同階段往往需要不同的生態因子或生態因子的不同強度。例如低溫對冬小麥的春化階段是必不可少的,但在其後的生長階段則是有害的。那些對生物的生長、發育、繁殖、數量和分佈起限制作用的關鍵性因子叫限制因子。有關生態因子(量)的限制作用有以下兩條定律。
生態系統
(2)謝爾福德耐受定理(Shelford’s law of tolerance)
生態學家V. E. Shelford於1913年研究指出,生物的生存需要依賴環境中的多種條件,而且生物有機體對環境因子的耐受性有一個上限和下限,任何因子不足或過多,接近或超過了某種生物的耐受限度,該種生物的生存就會受到影響,甚至滅絕。這就是Shelford耐受定律。後來的研究對Shelford耐受定律也進行了補充:每種生物對每個生態因子都有一個耐受範圍,耐受範圍有寬有窄;對所有因子耐受範圍都很寬的生物,一般分佈很廣;生物在整個發育過程中,耐受性不同,繁殖期通常是一個敏感期;在一個因子處在不適狀態時,對另一個因子的耐受能力可能下降;生物實際上並不在某一特定環境因子最適的範圍內生活,可能是因為有其他更重要的因子在起作用。
最小因子定律和耐受性定律的關係,可以從以下三個方面理解,首先,最小因子定律只考慮了因子量的過少,而耐受性定律既考慮了因子量的過少,也考慮了因子量的過多;其次,耐受性定律不僅估計了限制因子量的變化,而且估計了生物本身的耐受性問題。生物耐受性不僅隨種類不同,且在同一種內,耐受性也因年齡、季節、棲息地的不同而有差異;同時,耐受性定律允許生態因子之間的相互作用,如因子替換作用和因子補償作用。
生態因子作用的直接性和間接性
直接參與生物生理過程或參與新陳代謝的因子屬於直接因子,如光、溫、水、土壤養分等。例如光可以促進種子萌發。而那些通過影響直接因子而對生物作用的因子,屬於間接因子,如海拔,坡向,經、緯度等就是間接因子,他們對生物的作用不亞於直接因子。例如四川二郎山的東坡濕潤多雨,分佈類型為常綠闊葉林;而西坡空氣感乾熱、缺水,只能分佈耐旱的的灌草叢,同一山體由於坡向不同,導致植被類型各異。
實際上,任何生態因子都不是孤立地對植物起作用,植物受到的是多種生態因子的綜合影響。
多種生態因素形成一個整體對植物的生長和發育起作用,稱為綜合效應。生態因素往往是相互關聯的,如光照強度的增加會引起溫度的提高。生態因素之間還可能出現相互補償作用,如同樣的光合作用強度在一定的範圍內既可以出強光照和低的CO2濃度下,也可以出弱光照和高的CO2濃度下。也就是說,高的CO2濃度在一定條件下對光照的不足有補償作用。
每種植物所能適應的生態因子的範圍都有一定的限度,超過這個限度,植物的生長、發育和繁殖等一系列的生命活動就會受到影響,甚至引起死亡。如玉米生長發育所需要的溫度最低不能低於9.4℃,不能高於46.1℃。
所謂 最低量率就是在不可缺少的有效養分中,數量上接近於臨界最低的一個限制植物的產量。這一規律是由德國人李比希(Liebig)發現的。他在作物栽培實踐中觀察到:作物需要一定種類和數量的礦物養分,當某種礦物養分處於其臨界最低值時,它對作物的產量影響最大。
每一生態因子都是隨時間而變化的,植物適應的不是單個生態因子在某一特定時間的量,而是適應該因子隨時間變化的過程。植物生長發育的不同階段對生態因子的質和量的要求不同。如低溫對某些植物的春化階段來說是必需的生態條件,而這一階段過後,低溫會對植物造成傷害。
生態因子在空間上不是均質的。不同生態因子在空間分佈上的差異性直接影響到植物的空間分佈。隨著任一生態因子在空間上按順序增強或者減弱,不同生態類型的植物按順序排列的現象稱為 生態序列。如北京附近的一些水體,在水體中間水深較大的地方出現金魚藻(Ceratophyllum demersum)等挺水植物,水稍淺時出現荇菜(Nymphoides peltatum)等浮水植物,接近岸邊出現香蒲(Thypha spp.)等挺水植物,上岸后首先出現苔草等濕生植物,隨著地形部位的變化進一步出現中生植物甚至中旱生植物,這就是一個隨水分條件控制的生態序列。
不同種、同一種的不同品種甚至同一品種的不同植株,對於同一生態因子的響應並非絕對一致。

生態因子
① 構成維持生物代謝和繁殖所必需的營養物質和理化條件。這些理化條件也都表現為能量或物質,如日照、溫度、pH值、滲透壓等。
② 構成種種破壞力量。例如天敵、自然災害(超限的理化條件)及某些人類活動(濫墾濫牧、工業污染等)。
③ 僅僅作為信息,誘發生物的節律性反應。例如日照和溫度的晝夜或季節變化,能引起植物的萌發、生長、開花等階段變化和動物的冬眠、遷徙等周期活動。
生態因子作用的直接對象是生物個體,但通過生物間的交互作用會影響到群體。同種動物的集群活動可以增加取食和避敵能力。群落食物中某環節的增減,常導致連鎖反應,例如天氣變化造成蝗群增長及其相變,繼而導致遷飛,破壞遷入地的大片植被。
生態因子的作用與生物的適應性密切相關。對於溫度,各物種反應不同,有些物種能適應的溫度卻可能使另一些物種死亡。一般說,生物在不同發育階段的適應性也不大相同。環境在變,生物的適應性也隨之改變。一個物種可能通過生理過程適應一個新環境,當新舊環境差別太顯著時,可能需要較長時期的適應過程,引種馴化便屬此類。在生物發展史中,生態因子作為選擇因素淘汰掉不適應的物種。生態因子還可能直接誘發基因突變或重組,促進生物進化的進程。
生物有機體不是孤立生存的,在其生存環境中甚至其體內都有其他生物的存在,這些生物便構成了生物因子。生物與生物因子之間發生各種相互關係,這種相互關係既表種內個體之間,也存在於不同的種間。生物之間的相互關係,可以概括為八種類型(表1)。表1 生物之間相互關係類型
| 類型 | A | B | 特點 | 類型 | A | B | 特點 |
| 競爭 | - | - | 彼此互相抑制 | 共生 | + | + | 彼此有利,分開后不能生活 |
| 捕食 | + | - | A殺死或吃掉B | 互惠 | + | + | 彼此有利,分開能獨立生活 |
| 寄生 | + | - | A寄生於B,對B有害 | 偏利 | + | 對A有益,對B無影響 | |
| 中性 | 彼此互不影響 | 偏害 | - | 對A有害,對B無影響 |
生物因子主要有食物、捕食者、寄生物和病原微生物。與非生物因子相比,生物因子對生物的影響有以下特點:一般情況下,生物因子隻影響到種群中的某些個體;生物因子對生物種群的影響程度通常與種群的密度有關;生物因子在相互作用、相互制約中產生了協同進化;生物因子一般僅直接涉及兩個物種或與其鄰近密切相關物種之間的關係。
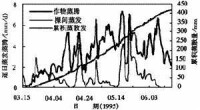
冬小麥限制因子分析
土壤是許多生物的棲息場所。土壤中的生物包括細菌、真菌、放線菌、藻類、原生動物、輪蟲、線蟲、蚯蚓、軟體動物、節肢動物和少數高等動物。土壤是生物進化的過渡環境。土壤中既有空氣,又有水分,正好成為生物進化過程中的過渡環境。土壤是植物生長的基質和營養庫。土壤提供了植物生活的空間、水分和必需的礦質元素。土壤是污染物轉化的重要場地。土壤中大量的微生物和小型動物,對污染物都具有分解能力。
2.土壤質地與結構對生物的影響
土壤是由固體、液體和氣體組成的三相系統,其中固體顆粒是組成土壤的物質基礎。土粒按直徑大小分為粗砂(2.0-0.2mm)、細粒(0.2-0.02mm)、粉砂(0.02-0.002mm)和粘粒(0.002mm以下)。這些大小不同的土粒的組合稱為土壤質地。根據土壤質地可把土壤分為砂土、壤土和粘土三大類。砂土的砂粒含量在50%以上,土壤疏鬆、保水保肥性差、通氣透水性強。壤土質地較均勻,粗粉粒含量高,通氣透水、保水保肥性能都較好,抗旱能力強,適宜生物生長。粘土的組成顆粒以細粘土為主,質地粘重,保水保肥能力較強,通氣透水性差。
土壤結構是指固體顆粒的排列方式、孔隙的數量和大小以及團聚體的大小和數量等。最重要的土壤結構是團粒結構(直徑0.25-10mm),團粒結構具有水穩定性,由其組成的土壤,能協調土壤中水分、空氣和營養物之間的關係,改善土壤的理化性質。
土壤質地與結構常常通過影響土壤的物理化學性質來影響生物的活動。

土壤因子調查
土壤溫度對植物種子的萌發和根系的生長、呼吸及吸收能力有直接影響,還通過限制養分的轉化來影響根系的生長活動。一般來說,低的土溫會降低根系的代謝和呼吸強度,抑制根系的生長,減弱其吸收作用;土溫過高則促使根系過早成熟,根部木質化加大,從而減少根系的吸收面積。
(2)土壤水分
土壤水分與鹽類組成的土壤溶液參與土壤中物質的轉化,促進有機物的分解與合成。土壤的礦質營養必需溶解在水中才能被植物吸收利用。土壤水分太少引起乾旱,太多又導致澇害,都對植物的生長不利。土壤水分還影響土壤內無脊椎動物的數量和分佈。
(3)土壤空氣
土壤空氣組成與大氣不同,土壤中O2的含量只有10-12%,在不良條件下,可以降至10%以下,這時就可能抑制植物根系的呼吸作用。土壤中CO2濃度則比大氣高几十到上千倍,植物光合作用所需的CO2有一半來自土壤。但是,當土壤中CO2含量過高時(如達到10-15%),根系的呼吸和吸收機能就會受阻,甚至會窒息死亡。
(4)土壤酸鹼度
土壤酸鹼度與土壤微生物活動、有機質的合成與分解、營養元素的轉化與釋放、微量元素的有效性、土壤保持養分的能力及生物生長等有密切關係。根據植物對土壤酸鹼度的適應範圍和要求,可把植物分成酸性土植物(pH<6.5)、中性土植物(pH6.5-7.5)和鹼性土植物(pH>7.5)。土壤酸鹼度對土棲動物也有類似影響。
基本信息
- 中文名
- 生態因子
- 外文名
- ecological factor