群落結構
群落結構
徠群落 (Biocoenosis)或稱為“生物群落”。生態學中,在一個群落生境內相互之間具有直接或間接關係的所有生物,或曰生物的總和。也有人作如下定義:生存在一起並與一定的生存條件相適應的動植物的總體。群
落生境是群落生物生活的空間。一個生態系統則是群落和群落生境的系統性相互作用。一個群落的生物物種佔據不同的小生境。一個群落的生物相互之間有著不同的關係。
1877年,德國動物學家Karl Möbius創造了biocenosis一詞(Möbius 1877),描述生活在一個生境(habitat)中相互作用的有機體,認為每一個生物群落都“支持著一定數量的有生命的生物”,強調在合適的條件下可能產生過量的後代,但由於空間和食物是有限的,“群落中個體總數不久又會回到它以前的適中狀態”,這使人回想起馬爾薩斯關於人口與它們受資源限制的理論。但在英語中biocenosis一詞很少使用,而常用Ecological community,指佔據同一地理區域的二個或以上不同種類的物種的集合。
生物群落作為一般名詞時與群落同義。生物群落一詞,只限於強調沿襲克列門茨和謝爾福德(F.E.Clements & V.E.Shelford,1939)的考慮方法。克列門茨認為,植物群落並不是個體和種的組合,而是用生長型(growth form)為代表的生態群組合,這種必須以生物個體為準據的新階段的有機體,稱之為複合生物(complex organism)。謝爾福德(1912)對動物曾使用了生理活動型這個名詞,它相當於植物的生長型,並稱之為生態種群(mores,用複數形,單數形為mos,涵義不同),它的組合即是動物群落。以後把植物和動物作為mune的機能生活形的集群,而進一步強調了統一性。演替是複合生物的發育、成長,相當於它的成體的頂極群落是僅僅由氣候決定的,也有把這種觀點特稱之為生物生態學(bio-ecology)的。
生物群落結構與生態系統的概念不同。後者不僅包括生物群落還包括群落所處的非生物環境,把二者作為一個由物質、能量和信息聯繫起來的整體。因此生物群落只相當於生態系統中的生物部分。
生物群落中的各種生物之間的關係主要有3類:
①營養關係,當一個種以另一個種,不論是活的還是它的死亡殘體,或它們生命活動的產物為食時,就產生了這種關係。又分直接的營養關係和間接的營養關係。採集花蜜的蜜蜂,吃動物糞便的糞蟲,這些動物與作為它們食物的生物種的關係是直接的營養關係;當兩個種為了同樣的食物而發生競爭時,它們之間就產生了間接的營養關係。因為這時一個種的活動會影響另一個種的取食。
②成境關係,一個種的生命活動使另一個種的居住條件發生改變。植物在這方面起的作用特別大。林冠下的灌木、草類和地被以及所有動物棲居者都處於較均一的溫度、較高的空氣濕度和較微弱的光照等條件下。植物還以各種不同性質的分泌物(氣體的和液體的)影響周圍的其他生物。一個種還可以為另一個種提供住所,例如,動物的體內寄生或巢穴共棲現象,樹木干枝上的附生植物等。
③助布關係,指一個種參與另一個種的分佈,在這方面動物起主要作用。它們可以攜帶植物的種子、孢子、花粉,幫助植物散布。
營養關係和成境關係在生物群落中具有最大的意義,是生物群落存在的基礎。正是這兩種相互關係把不同種的生物聚集在一起,把它們結合成不同規模的相對穩定的群落。
1.具有一定的動態特徵
2.具有一定的邊界特徵
3.具有一定的分佈範圍
4.具有一定的結構
5.具有一定的種類組成
6.具有一定的群落環境
7.不同物種之間的相互影響
8.各物種不具有同等的群落學重要性
⒈一個群落環境的小環境越是多樣,那麼其中的群落就越是物種豐富。(熱帶雨林)
⒉一個群落環境的非生物因素和全球平均值相差越大,其物種也會越少,但個體數目卻會越多。這個區域的群落通常是高特異性的。(鹽湖,深海)
⒊一個群落環境的生存環境越是緩慢連續的交替改變,群落也會越是豐富。(珊瑚礁)
在生物群落中,各個種群佔據了不同的空間,使群落具有一定的結構。群落的結構包括垂直結構和水平結構。
相對穩定了的生物群落其重要特徵之一,是具有一定的空間結構。群落中各種生物在空間上的配置狀況,即為群落的結構。群落結構包括形態方面的結構和生態方面的結構,前者包括垂直結構和水平結構,後者指層片結構。
包徠括空間結構、時間組配和種類結構。
不同生活型的植物(喬木、灌木、草本)生活在一起,它們的營養器官配置在不同高度(或水中不同深度),因而形成分層現象。分層使單位面積上可容納的生物數目加大,使它們能更完全、更多方面地利用環境條件,大大減弱它們之間競爭的強度;而且多層群落比單層群落有較大的生產力。
分層現象在溫帶森林中表現最為明顯,例如溫帶落葉闊葉林可清晰地分為喬木、灌木、草本和苔蘚地衣(地被)4層。熱帶森林的層次結構最為複雜,可能有的層次最為發育,特別是喬木層,各種高度的巨樹、一般樹和小樹密集在一起,但灌木層和草本層常常不很發育。草本群落一樣地分層,儘管層次少些(通常只分為草本層和地被層)。
群落不僅地上分層,地下根系的分佈也是分層的。群落地下分層和地上分層一般是相應的;喬木根系伸入土壤的最深層,灌木根系分佈較淺,草本植物根系則多集中土壤的表層,蘚類的假根則直接分佈在地表。
生物群落結構的垂直分層與光照條件密切相關,每一層的植物適應於該層的光照水平,並降低下層的光強度。在森林中光強度向下遞減的現象最為明顯。最上層樹處於全光照之中,平均說來,到達下層小樹的光只有上層樹(全光照)的10~50%,灌木層只有5~10%,而草本層則只剩1~5%了。隨著光照強度的變化,溫度、空氣濕度也發生變化。
每一層植物和被它們所制約的小氣候為生活於其中的特有動物創造一定的環境,因此動物在種類上也表現出分層現象,不同的種類出現於不同層次,甚至同一種的雌雄個體,也分佈於不同的層次。例如,在森林中可以區分出3組鳥種:在樹冠中採食的,接近地面的,以及生活在其間的灌木和矮樹簇葉中的。
林地也由於枯枝落葉層的積累和植物對土壤的改造作用,創造了特殊的動物棲居環境。較高的層(草群,下木)為吃植物的昆蟲、鳥類、哺乳動物和其他動物所佔據。在枯枝落葉層中,在腐爛分解的植物殘體、蘚類、地衣和真菌中,生活著昆蟲、蜱、蜘蛛和大量的微生物。在土壤上層,擠滿了植物的根,這裡居住著細菌、真菌、昆蟲、蜱、蠕蟲。有時在土壤的某種深度還有穴居的動物。
當然,也存在一些層外生物,它們不固定於某一個層。例如藤本植物、附生植物,以及從一個層到另一個層自由活動的動物。它們使劃分層次困難化;在結構極其複雜的熱帶雨林中經常見到這種情況。
因為下層生物是在上層植物遮蔭所形成的環境中發育起來的,所以生物群落中不同層的物種間有密切的相互作用和相互依賴關係。群落上層植物強烈繁生,相應地下層植物的密度就會降低;而如果由於某種原因上層植物變得稀疏,下層的光照、熱等狀況得到改善,同時土壤中礦物養分因釋放加強而增高,下層植物發育便會加強。下層的繁茂生長也對動物棲居者有利。這種情況特別反映在森林群落中,哪裡喬木層稀疏便會導致那裡的灌木或喜光草本植被的豐富繁生。而喬木層的完全鬱閉,有時甚至抑制最耐蔭的草本和蘚類。
生物群落不僅有垂直方向的結構分化,而且還有水平方向的結構分化。群落在水平方向的不均勻性表現為以斑塊出現;在不同的斑塊上,植物種類、它們的數量比例、鬱閉度、生產力以及其他性質都有不同。例如在一個草原地段,密叢草針茅是最佔優勢的種類,但它並不構成連續的植被,而是彼此相隔一定的距離(30~40厘米)分佈的。各個針茅草叢之間的空間,則由各種不同的較小的禾本科植物和雙子葉雜類草佔據著,並混有鱗莖植物。但其中的某些植物也出現於針茅草叢的內部。因此,伴生少數其他植物的針茅草叢同針茅草叢之間生長有其他草類的空隙,它們在外貌、在種間數量關係和質量關係上都有很明顯的不同。但它們的差別與整個植物群落(針茅草原)比較起來,是次一級的差別,而且是不很明顯的和不穩定的。在森林中,在較陰暗的地點和較明亮的地點,也可以觀察到在植物種類的組成和數量比例方面以及其他方面的類似差異。群落內水平方向上的這種不一致性,叫做群落的鑲嵌性。這種不一致性在某些情況下是由群落內環境的差別引起的,如影響植物種分佈的光強度不同或地表有小起伏;在某些情況下是由於共同親本的地下莖散布形成的植物集群所引起;在另外的情況下,它們可能由種之間的相互作用引起,例如在寄主種的根出現的地方形成斑塊狀的寄生植物。動物的活動有時也是引起不均一性的原因。植物體通常不是隨機地散佈於群落的水平空間,它們表現出成叢或成簇分佈。許多動物種群,不論在陸地群落或水生群落,也具有成簇分佈的性質。相比之下,有規則的分佈是比較不常見的。某些荒漠中灌木的分佈、鳴禽和少數其他動物的均勻分佈是這種有規則分佈的例子。
1.垂直結構
群落的垂直結構指群落在垂直方面的配置狀態,其最顯著的特徵是成層現象,即在垂直方向分成許多層次的現象。
群落的成層性包括地上成層和地下成層。層的分化主要決定於植物的生活型,生活型不同,植物在空中占居的高度以及在土壤中到達的深度就不同,水生群落則在水面以下不同深度形成物種的分層排列,這樣就出現了群落中植物按高度(或深度)配置的成層現象。
成層現象在森林群落表現最為明顯,而以溫帶闊葉林和針葉林的分層最為典型,熱帶森林的成層結構則最為複雜。一般按生長型把森林群落從頂部到底部劃分為喬木層、灌木層、草本層和地被層(苔蘚地衣)四個基本層次,在各層中又按植株的高度劃分亞層,例如熱帶雨林的喬木層通常分為三個亞層。草本群落則通常只有草本層和地被層。
在層次劃分時,將喬木和其他生活型植物不同高度的幼苗划入實際所逗留的層中,生活在各層中的地衣、藻類、藤本等層間植物通常也歸入相應的層中。
群落的地下分層和地上分層一般是相應的。森林群落中的喬木根係為分佈到土壤的深層,灌木根系較淺,草本植物的根系則大多分佈在土壤的表層。草本群落的地下分層比地上分層更為複雜。
群落的成層性保證了植物在單位空間中更充分利用自然環境條件。如在發育成熟的森林中,上層喬木可以充分利用陽光,而林冠下為那些能有效利用弱光的下木所佔居,林下灌木層和草本層能夠利用更微弱的光線、草本層往下還有更耐蔭的苔蘚層。
生物群落中動物的分層現象也很普遍。動物分層主要與食物有關,其次還與不同層次的微氣候條件有關。如東歐亞大陸北方針葉林區,在地被層和草本層中,棲息著兩棲類、爬行類、鳥類(丘鷸、楱雞)、獸類(黃鼠)和各種鼠形嚙齒類;在森林的下層——灌木林和幼林中,棲息著鶯、葦鶯和花鼠等;在森林的中層棲息著山雀、啄木鳥、松鼠和貂等,而在樹冠層則棲息著柳鶯、交嘴和戴菊等。
水域中某些水生動物也有分層現象,這主要決定於陽光、溫度、食物和含氧量等。比如湖泊,在一年當中湖水沒有循環流動的時候,浮遊動物都表現出明顯的垂直分層現象,它們多分佈在較深的水層,在夜間則上升到表層來活動,這是因為浮遊動物一般都是趨向弱光的。
2.水平結構
群落的水平結構指群落的水平配置狀況或水平格局,其主要表現特徵是鑲嵌性。
鑲嵌性即植物種類在水平方向不均勻配置,使群落在外形上表現為斑塊相間的現象。具有這種特徵的群落叫做鑲嵌群落。在鑲嵌群落中,每一個斑塊就是一個小群落,小群落具有一定的種類成份和生活型組成,它們是整個群落的一小部分。例如,在森林中,林下陰暗的地點有一些植物種類形成小型的組合,而在林下較明亮的地點是另外一些植物種類形成的組合。這些小型的植物組合就是小群落。內蒙古草原上錦雞兒灌叢化草原是鑲嵌群落的典型例子。在這些群落中往往形成1~5米左右的錦雞兒叢,呈園形或半園形的丘阜。這些錦雞兒小群落內部由於聚集細土、枯枝落葉和雪,因為具有良好的水分和養分條件,形成一個局部優越的小環境。小群落內部的植物較周圍環境中返青早,生長發育好,有時還可以遇到一些越帶分佈的植物。
群落鑲嵌性形成的原因,主要是群落內部環境因子的不均勻性,例如小地形和微地形的變化,土壤溫度和鹽漬化程度的差異,光照的強弱以及人與動物的影響。在群落範圍內,由於存在不大的低地和高地因而發生環境的改變形成鑲嵌,這是環境因子的不均勻性引起鑲嵌性的例子。由於土中動物,例如田鼠活動的結果,在田鼠穴附近經常形成不同於周圍植被的斑塊,這是動物影響鑲嵌性的例子。
3.層片結構
層片一詞系瑞典植物學家加姆斯發明創造。他起初賦於這一概念以三個方面的內容,即把層片劃分為三級:一級層片,即同種個體的組合;二級層片,即同一生活型的不同植物的組合;三級層片,即不同生活型的不同種類植物的組合。一般群落學研究中使用的層片概念,均相當於加姆斯的二級層片,即每一個層片都是由同一生活型的植物所組成。
生活型是植物對外界環境適應的外部表現形式,同一生活型的植物不但體態上是相似的,而且在形態結構、形成條件、甚至某些生理過程也具相似性。如今廣泛採用的生活型劃分是郎基耶爾的系統。他按照休眠芽在不良季節的著生位置把植物的生活型分成五大類群,高位芽植物(25厘米以上),地上芽植物(25厘米以下)、地面芽植物(位於近地面土層內)、隱芽植物(位於較深土層或水中)和一年生植物(以種子越冬),在各類群之下再細分為30個較小的類群。我國植被學著作中採用的是按體態劃分的生活型系統,該系統把植物分成木本植物、半木本植物、草本植物、葉狀體植物四大類別,再進一步劃分成更小的或低級的單位。對於層片的劃分,可以根據研究的需要,分別使用上述系統中的高級劃分單位或低級單位。
層片作為群落的結構單元,是在群落產生和發展過程中逐步形成的。它的特點是具有一定的種類組成,它所包含的種具有一定的生態生物學一致性,並且具有一定的小環境,這種小環境是構成植物群落環境的一部分。
需要說明一下層片與層的關係問題。在概念上層片的劃分強調了群落的生態學方面,而層次的劃分,著重於群落的形態。層片有時和層是一致的,有時則不一致。例如分佈在大興安嶺的興安落葉松純林,興安落葉松組成喬木層,它同時也是該群落的落葉針葉喬木層片。在混交林中,喬木層是一個層,但它由闊葉樹種層片和針葉樹種層片兩個層片構成。在實踐中,層片的劃分比層的劃分更為重要,但劃分層次往往是區分和分析層片的第一步。
和層結構一樣,群落層片結構的複雜性,保證了植物全面利用生境資源的可能性,並且能最大程度地影響環境,對環境進行生物學改造。
組成群落的生物種在時間上也常表現出“分化”,即在時間上相互“補充”,如在溫帶具有不同溫度和水分需要的種組合在一起:一部分生長於較冷季節(春秋),一部分出現於炎熱季節(夏)。例如,在落葉闊葉林中,一些草本植物在春季樹木出葉之前就開花,另一些則在晚春、夏季或秋季開花。隨著不同植物出葉和開花期的交替,相聯繫的昆蟲種也依次更替著:一些在早春出現,另一些在夏季出現。鳥類對季節的不同反應,表現為候鳥的季節性遷徙。生物也表現出與每日時間相關的行為節律:一些動物白天活動;另一些黃昏時活動;還有一些在夜間活動,白天則隱藏在某種隱蔽所中。大多數植物種的花在白天開放,與傳粉昆蟲的活動相符合;少數植物在夜間開花,由夜間動物授粉。許多浮遊動物在夜間移向水面,而在白天則沉至深處遠離強光,但是不同的種具有不同的垂直移動模式和範圍、潮汐的複雜節律控制著許多海岸生物的活動。土壤棲居者也有晝夜垂直移動的種類。
每一個具體的生物群落以一定的種類組成為其特徵。但是不同生物群落種類的數目差別很大。例如,在熱帶森林的生物群落中,植物種以萬計,無脊椎動物種以10萬計,脊椎動物種以千計,其中的各個種群間存在非常複雜的聯繫。
生物群落
凍原和荒漠群落的種數要少得多。根據蘇聯學者Б.А.季霍米羅夫的資料,在西伯利亞北部的泰梅爾半島的凍原生物群落中共有139種高等植物,670種低等植物,大約1000種動物和2500種微生物。與此相應。這些生物群落的生物量和生產力,也比熱帶森林小得多。
生物群落中生物的複雜程度用物種多樣性這一概念表示。多樣性與出現於某一地區的生物種的數量有關,也與個體在種之間的分佈的均勻性有關。例如,兩個群落都含有5個種和100個個體,在一個群落中這100個個體平均地分配在全部5個種之中,即每1個種有20個個體,而在另一個群落中80個個體屬於1個種,其餘20個個體則分配給另外的4個種,在這種情況下,前一群落比后一群落的多樣性大。
在溫帶和極地地區,只有少數物種很常見,而其餘大多數物種的個體很稀少,它們的種類多樣性就很低;在熱帶,個體比較均勻地分佈在所有種之間,相鄰兩棵樹很少是屬於同種的(熱帶雨林),種類多樣性就相對較高。群落的種類多樣性決定於進化時間、環境的穩定性以及生態條件的有利性。熱帶最古老,形成以來環境最穩定,高溫多雨氣候對生物的生長最為有利,以生物群落的種類多樣性最大。在嚴酷的凍原環境中,情況相反,所以種類多樣性低。
每種植物在群落中所起的作用是不一樣的。常常一些種以大量的個體,即大的種群出現;而另一些種以少量的個體,即小的種群出現。個體多而且體積較大(生物量大)的植物種決定了群落的外貌。例如,絕大多數森林和草原生物群落的一般外貌決定於一個或若干個植物種,如中國山東半島的大多數櫟林決定於麻櫟,燕山南麓的松林決定於油松,內蒙古高原中東部錫盟的針茅草原決定於大針茅或克氏針茅等。在由數十種甚至百餘種植物組成的森林中,常常只有一種或兩種喬木提供90%的木材。群落中的這些個體數量和生物量很大的種叫做優勢種,它們在生物群落中占居優勢地位。優勢種常常不止一個,優勢種中的最優勢者叫建群種,通常陸地生物群落根據建群植物種命名,例如,落葉闊葉林、針茅草原、泥炭蘚沼澤等。建群種是群落的創建者,是為群落中其他種的生活創造條件的種。例如,雲杉在泰加帶形成稠密的暗針葉林,在它的林冠下,只有適應於強烈遮蔭條件,高的空氣濕度和酸性灰化土條件的植物能夠生活;相應於這些因素,在雲杉林中還形成特有的動物棲居者。因此在該情況下雲杉起著強有力的建群種的作用。
松林中的建群種是松樹,但與雲杉相比,它是較弱的建群種,因為松林樹榦稀疏,樹冠比較不密接,比較透光,它的植物和動物的種類組成遠比雲杉林豐富和多樣。在松林中甚至見到能生活在林外環境中的植物。
溫帶和寒帶地區的生物群落中,建群種比較明顯;無論森林群落、灌木群落、草本群落或蘚類群落,都可以確定出建群種(有時不止一個)。亞熱帶和熱帶,特別是熱帶的生物群落,優勢種不明顯,很難確定出建群種來。除優勢種外,個體數量和生物量雖不佔優勢但仍分佈廣泛的種是常見種;個體數量極少,只偶爾出現的種是偶見種。
生物群落中的大多數生物種,在某種程度上與優勢種和建群種相聯繫,它們在生物群落內部共同形成一個物種的綜合體,叫做同生群。同生群也是生物群落中的結構單位。例如一個優勢種植物,和與它相聯繫的附生、寄生、共生的生物以及以它為食的昆蟲和哺乳動物等共同組成一個同生群。
生活在一個群落中的多種多樣的生物種,是在長期進化過程中被選擇出來能夠在該環境中共同生存的種。它們中每一個佔據著獨特的小生境,並且在改造環境條件、利用環境資源方面起著獨特的作用。群落中每一個生物種所佔據的特定的生境和它執行的獨特的功能的結合,叫做生態位。因此,一個生物群落的物種多樣性越高,其中生態位分化的程度也越高。
群落結構總體上是對環境條件的生態適應,但在其形成過程中,生物因素起著重要作用,其中作用最大的是競爭與捕食。
競爭對群落結構的影響
競爭對群落結構的形成有重要影響。對種間競爭在形成群落結構的作用問題上,最直接的證據可能是在自然群落中對物種進行引進或去除實驗,Schoner和 Cornell 就分別對種間競爭研究的文獻進行過統計(分別達184和72 例研究),平均有90%的例證說明有種間競爭,表明自然群落中競爭是相當普遍的。他們的結果還表明,海洋生物中有種間競爭的比例較陸地生物多;大型生物間比小型生物間高;而植食性昆蟲中競爭比例低,因為綠色植物到處都有,較豐富,很少被一食而空,所以為食物資源而競爭的可能性比較小。
已有證據表明,競爭是群落形成的一個重要驅動因素。但競爭的重要性在多個群落間顯然是不同的,而且常常只是影響物種之間相互作用的一小部分。
那麼為什麼許多調查結果顯示的競爭往往是不強烈的呢?
一般認為:① 自然選擇可能已有效地通過生態位劃分而避免了競爭(或者抹去了過去競爭的痕迹);② 在一個環境斑塊中,具有強競爭力的物種共存,因為它們並不利用相同的資源;③ 物種也許僅僅在種群爆發、資源短缺時才發生競爭。
捕食對群落結構的影響
捕食對形成群落結構的影響,視捕食者是泛化種還是特化種而不同。對泛化種來說,捕食使種間競爭緩和,並促進多樣性提高。但當取食強度過高時,物種數亦隨之降低。對特化種來說,隨被選食的物種是優勢種還是劣勢種而異。如果被選擇的是優勢種,則捕食能提高多樣性,如果捕食者喜食的食競爭力弱的劣勢種,多樣性就會隨著呈現型下降趨勢。
干擾對群落結構的影響
干擾是自然界的普遍現象。干擾不同於災難,不會產生巨大的破壞作用,但它經常會反覆的出現,使物種沒有充足的時間進化。近代多數生態學家認為干擾是一種有益的生態現象,它引起群落的非平衡特性,強調了干擾在群落結構形成和動態中的作用。
① 干擾與群落的缺口
連續的群落中出現缺口是非常普遍的現象,而缺口經常由干擾構成。森林中的缺口可能由大風、雷電、砍伐、火燒等引起;草地群落的干擾包括放牧、動物挖掘、踐踏等。干擾造成群落的缺口以後,有的在沒有繼續干擾的條件下會逐漸地恢復,但缺口也可能被周圍群落的任何一個種侵入和佔有,並發展為優勢者,哪一種是優勝者完全取決於隨機因素,這可稱為對缺口的抽彩式競爭。
但是,有些群落所形成的缺口,其物種更替是有規律性的。新打開的缺口常常被擴散能力強的一個或幾個先鋒種所入侵。由於它們的活動,改變了條件,促進了演替中期種的入侵,最後為頂級種所替代。在這種情況下,多樣性開始較低,演替中期增加,但到頂級期往往稍有降低。
② 中度干擾假說
缺口形成的頻率影響物種多樣性,據此,T.W.Connell等提出了中度干擾假說,即中等程度的干擾水平能維持高多樣性。其理由是:
一、在一次干擾后少數先鋒物種入侵缺口,如果幹擾頻繁,則先鋒種不能發展到演替中期,因而多樣性較低;
二、如果幹擾間隔期很長,使演替過程能發展到頂級期,多樣性也不很高;
三、只有中等干擾程度使多樣性維持最高水平,它允許更多的物種入侵和定居。
| 類 型 | 作用方式 | 實 例 |
| 捕食者 | 抑制競爭者 | 海洋:海獺、海膽陸地:依大小選擇性採食種子的動物 |
| 食草動物 | 抑制競爭者 | 大象、兔子 |
| 病原體和寄生物 | 抑制捕食者、食草動物競爭者 | 粘液瘤菌、采采蠅 |
| 競爭者 | 抑制競爭者 | 演替中的物種更替,如森林中的優勢樹種和雜草 |
| 共生種 | 有效的繁殖 | 關鍵的共生種依賴的植物資源傳粉者、傳播者 |
| 掘土者 | 物理干擾 | 兔子、地鼠、白蟻、河狸、河馬 |
| 系統過程調控者 | 影響養分傳輸速率 | 固氮菌、菌根真菌分解者 |
群落的環境不是均勻一致的,空間異質性的程度越高,意味著有更加多樣的小生境,所以能允許更多的物種共存。
植物群落研究中大量資料說明,在土壤和地形變化豐富的地方,群落會有更多的物種;而平坦、同質土壤的群落多樣性低。Harman 研究了淡水系統軟體動物種數與空間異質性的相關性,也得出棲境質類型越多,淡水軟體動物種數越多的正相關的關係結果。
MacArthur等通過研究鳥類多樣性與植物物種多樣性和取食多樣性之間的關係,發現鳥類多樣性與植物種數的相關,不如與取食——高度多樣性相關明顯。所以對於鳥類生活來說,植被的分層結構比物種組成更為重要。在草地和灌叢群落中,垂直結構不如森林群落明顯,而水平結構,即鑲嵌性和斑塊性,就可能起到決定性作用。
從表上可知,每一類植物群落都是由幾種生活型的植物所組成,但其中有一類生活型佔優勢,生活型與環境關係密切,高位芽植物佔優勢是溫暖、潮濕氣候地區群落的特徵,如熱帶雨林群落;地面芽植物佔優勢的群落,反映了該地區具有較長的嚴寒季節,如溫帶針葉林、落葉林群落;地下芽植物佔優勢,反映了該地區環境比較濕冷,如長白山寒溫帶暗針葉林;一年生植物佔優勢則是乾旱氣候的荒漠和草原地區群落的特徵,如東北溫帶草原。
| 群落類型 | Ph. | Ch. | H. | Cr. | T. |
| 西雙版納熱帶雨林 | 94.7 | 5.3 | |||
| 鼎湖山南亞熱帶常綠闊葉林 | 84.5 | 5.4 | 4.1 | 4.1 | |
| 浙江中亞熱帶常綠闊葉林 | 76.7 | 1.0 | 13.1 | 7.8 | 2 |
| 秦嶺北坡溫帶落葉闊葉林 | 52.0 | 5.0 | 38.0 | 3.7 | 1.3 |
| 長白山寒溫帶暗針葉林 | 25.4 | 4.4 | 39.8 | 26.4 | 3.2 |
| 東北溫帶草原 | 3.6 | 2.0 | 41.1 | 19.0 | 33.4 |
島嶼的物種數與面積的關係
島嶼由於與大陸隔離,生物種遷入和遷出的強度低於周圍連續的大陸。許多研究證實,島嶼中的物種數目與島嶼的面積有密切關係。島嶼面積越大,島嶼上的物種數越多,島嶼面積與島嶼上物種數的關係可以用簡單方程描述為:
S = cAz
或用對數表示:lgS = lgc + z (lgA)
式中:S —— 種數;
A —— 面積;
z 、c —— 兩個常數。
⒉3.4.2 MacArthur 的平衡說
島嶼上的物種數雖決定於島嶼的面積,但它是物種遷入、遷出和滅亡平衡的結果,這是一種動態平衡,不斷地有物種滅亡,也不斷地由同種或別種的遷入而補償滅亡的物種。
島嶼上物種的平衡關係可用圖3 — 2說明,以遷入率曲線為例,當島上無留居種時,任何遷入個體都是新的,因而遷入率高。隨著留居種數加大,種的遷入率就下降。當種源庫(大陸上的種)所有的種在島上都有時,遷入率為零。滅亡率則相反,留居種數越多,滅亡率越高。遷入率取決於島嶼與大陸距離的遠近和島嶼的大小,離大陸較近且面積大的島,其遷入率高;遠而小的島,遷入率低。同樣,滅亡率也受島的大小影響。
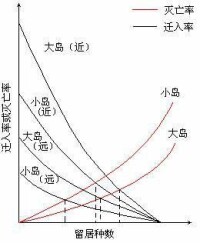
圖 3 —2 不同島上物種遷入率和滅亡率
① 島嶼上的物種數不隨時間而變化;
② 這是一種動態平衡,即滅亡種不斷地被新遷入的種所代替;
③ 大島比小島能“供養”更多的種;
④ 隨島距大陸的距離由近到遠,平衡點的種數逐漸降低。
平衡學說和非平衡學說
對於形成群落結構的一般理論,有兩種對立的觀點,即平衡學說和非平衡學說。
平衡學說認為共同生活在同一群落中的物種種群處於一種穩定狀態。其中心思想是:
①共同生活的物種通過競爭、捕食和互利共生等種間相互作用而互相牽制;
②生物群落具有全局穩定性特點,種間相互作用導致群落的穩定特性,在穩定狀態下群落的物種組成和各種群落數量都變化不大;
③群落實際上出現的變化是由於環境的變化,即所謂的干擾所造成的,並且干擾是逐漸衰亡的。因此,平衡學說把生物群落視為存在於不斷變化著的物理環境中的穩定實體。
非平衡學說的主要依據就是中度干擾理論。該學說認為,構成群落的物種始終處於變化之中,群落不能達到平衡狀態,自然界的群落不存在全局穩定性,有的只是群落的抵抗性(群落抵抗外界干擾的能力)和恢復性(群落在受干擾后恢復到原來狀態的能力)。
比較平衡學說和非平衡學說,除對干擾的作用強調不同以外,一個基本的區別是:平衡學說的注意焦點是系統處於平衡點的性質,而對於時間和變異性注意不足;而非平衡學說則把注意的焦點是系統在平衡點周圍的行為變化過程,特彆強調時間和變異性。兩個學說另一重要區別是把群落視為封閉系統還是開放系統。
可從生產力、有機物質的分解和養分循環3方面來描述。
群落中的綠色植物通過光合作用從無機物質製造有機化合物,這是生物群落的最重要的功能。在光合作用過程中,一段時間內由植物生產的有機物質的總量叫總初級生產力,通常以克/米2·年或千卡/米2·年表示。可是植物為了維持生存要進行呼吸作用,呼吸作用要消耗一部分光合作用生成的有機物質,剩餘的部分才用於積累(生長);一段時間內植物在呼吸之後餘下的有機物質的數量,叫凈初級生產力,例如在森林中,60~75%的總生產量可能被植物呼吸掉,餘下的40~25%才是凈生產量,在水生群落中不到總生產量的一半可能被植物呼吸掉。凈初級生產量隨時間的前進會逐漸積累,日益增多,到任一觀測時間為止積累下來的數量就是植物生物量。生物量以克/米2或千克/公頃表示。
生態學上更關心的是群落的生產力,即單位時間內的生產量。對於陸地或水底群落,是計算單位面積內的生物量數量,而對於浮遊和土壤群落則按單位容積確定。因而生物生產力乃是平方米面積上(或立方米容積中)在單位時間內的生產量,經常以碳的克數或干有機物質的克數表示。
生物生產力不能與生物量混淆。例如,一年內單位面積上的浮遊藻類合成的有機物質可能和高生產力的森林一樣多,但因大部分被異養生物所消費,故前者的生物量只有後者的十萬分之一。按照生產力,草甸草原的生物量年增長量比針葉林的大得多。根據蘇聯的資料,在中等草甸草原植物生物量為每公頃23噸的情況下,它們的年生產量達到每公頃10噸,而在針葉林,在植物生物量為每公頃200噸的情況下,年增長量每公傾只有6噸。小型哺乳動物比大型哺乳動物有較大的生長和繁殖速度,在相等的生物量的情況下提供較高的生產量。
消耗初級生產量的消費者也形成自己的生物量。它們在一段時間內的有機物質生產量叫次級生產量,即異養生物的生產量。消費者形成產量的速率叫次級生產力。
表1
地球上不同群落的初級生產力差別很大。R.H.惠特克按初級生產力將地球生物群落分為4類(見表1)。綠色植物的生物生產量,一部分以枯枝落葉的形式被分解者分解,一部分被風、水或其他動力帶至群落之外,一部分沿食物鏈傳遞。餘下的部分以有機物質的形式積累在群落中。
在許多群落中,動物從活植物組織得到的凈初級生產量部分要比植物組織死亡之後被分解者細菌和真菌等利用的部分小得多。在森林中,動物食用的大約不到葉組織的10%,不到活木質組織的1%,大部分落到地面形成覆蓋土壤表面的枯枝落葉層,被各種各樣的土壤生物所利用。這些土壤生物包括吃死植物組織和死動物組織的食腐者,分解有機質的細菌和真菌,以及以這些生物為食的動物。雖然動物有助於枯枝落葉的破壞,但細菌和真菌在把死有機物質還原成無機最終產物方面起最主要的作用。
分解者的生物量與消費者的相比是很小的,與生產者的相比更小。然而,物質量微小的分解者的活動在群落功能中十分重要。群落中全部死亡生物的殘體依賴分解者進行破壞。如果沒有分解者的分解活動,生物的死亡殘體將不斷地積累,像在酸沼中形成泥炭那樣。不僅群落的生產力可能由於養分被閉鎖在死組織中而受限制,而且整個群落也將不能存在。
群落中生產者從土壤或水中吸收無機養分,如氮、磷、硫、鈣、鉀、鎂以及其他元素,利用這些元素合成某些有機化合物,組成原生質和保持細胞執行功能。消費者動物從吃植物或其他動物取得這些元素。分解者在分解動、植物廢物產品和死亡殘體時,養分又釋放歸還到環境中,再被植物吸收。這便是養分循環,或稱物質的生物性循環。例如,在森林中,某種養分從土壤被吸收進入樹根,通過樹的輸導組織向上運輸到葉子,這時可能被吃葉子的蠋所食入,然後又被吃蠋的鳥所利用,直到鳥死亡后,被分解釋放歸還到土壤,再被植物根重新吸收。許多養分採取較短的途徑從森林樹木回到土壤──隨植物組織掉落到枯枝落葉層而被分解,或者在雨水淋洗下由植物表面落到土壤。
不同群落參加循環的養分數量和循環的速度不同。在一部分群落中,某些元素的較大部分保持在植物組織中,只有較小部分在土壤和水中遊離。例如溶於水中的磷酸鹽數量與浮游生物細胞和顆粒中的數量比較起來只是小部分。在熱帶森林,大部分養分保持在植物組織中,被雨水淋洗到土壤的養分和枯枝落葉腐敗分解時釋放出的養分,很快被重新吸收。但當一片森林被採伐或火燒后,通過侵蝕和養分在土壤水中的向下移動,造成養分的大量損失。在開闊大洋中隨著浮游生物細胞和有機顆粒的下沉,養分也被攜帶到深處,因而在進行光合作用的光亮表層水中養分很少,所以生產力很低。
群落結構在生態農業上有廣泛應用,如:
1.在稻田裡養鴨
2.用加工后的雞糞餵豬
3.利用玉米螟蟲天敵赤眼蜂防治蟲害
4.坡上種果樹 樹下養家禽
5.稻藕魚立體生態農業
基本信息
- 中文名
- 群落結構
- 外文名
- Biocoenosis
- 出處
- 1877年,德國動物學家Karl Möbius創造了biocenosis一詞(Möbius 1877),描述生活在一個生境(habitat)中相互作用的有機體,認為每一個生物群落都“支持著一定數量的有生命的生物”
- 提出者
- 德國動物學家Karl Möbius
- 詞性
- 名詞
- 分類
- 結構