系統發生樹
描述物種之間進化關係的樹
系統發生樹(英文:Phylogenetic tree)又稱為演化樹(evolutionary tree),是表明被認為具有共同祖先的各物種間演化關係的樹。是一種親緣分支分類方法(cladogram)。在樹中,每個節點代表其各分支的最近共同祖先,而節點間的線段長度對應演化距離(如估計的演化時間)。
系統發生樹(英文:phylogenetic tree或evolutionary tree)是表明被認為具有共同祖先的各物種相互間演化關係的樹,又被譯作系統發育樹、系統演化樹、系統進化樹、種系發生樹、演化樹、進化樹、系統樹。它用來表示系統發生研究的結果,用它描述物種之間的進化關係。
根據有根和無根來區分:
樹可分為有根樹和無根樹兩類。有根樹是具有方向的樹,

根據系統發生樹可推斷出物種的起源
基因樹和物種樹:
基於單個同源基因差異構建的系統發生樹應稱之為基因樹。因為這種樹代表的僅僅是單個基因的進化歷史。而不是它所在物種的進化歷史。物種樹一般最好是從多個基因數據的分析中得到。例如一項關於植物進化的研究中,用了100個不同的基因來構建物種樹,因為進化是發生在生物體種群水平上的,而不是發生在個體水平上的,雖然表面上不需要更多的數據,但實際上還是有必要的。基因樹和物種樹之間的差異是很重要的,如果只用等位基因來構建物種數,那許多人和大猩猩就會分到一起,而不是和其他人分到一起。
系統樹是一種分支圖(英文cladogram)。在樹中,每個節點代表其各分支的近共同祖先,而節點間的線段長度對應演化距離(如估計的演化時間)。
系統發生樹有時也稱系統樹圖,它是由一系列節點和分支組成的。其中每一個節點代表一個分類單元,分支末端的節點對應 一個基因或者生物體。與外部節點對應,內部節點代表一個推斷出的共同祖先。系統發生樹結構的基本信息在計算機程序中常常用一組嵌套的圓括弧表示,成為newick格式。
利用SSU rRNA繪製的系統演化樹,三個最大分支(域)分別為細菌、古菌和真核生物。
非加權分組平均法:UPGAM(Unweighted pair group method with arithmetic mean)
矩陣法:鄰接法 neighbor-joining (NJ)
簡約法:最大簡約法 maximum parsimony (MP)
似然法:最大似然法 maximum likelihood (ML)
后驗概率法:貝葉斯法 Bayesian
要構建一個進化樹(phyligenetic tree)。構建進化樹的演演算法主
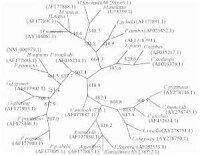
基因的系統發生樹
用於構建系統發生樹的數據分為兩類:特徵數據(存在有限不同狀態的特徵)和距離數據(兩個數據之間所有兩兩差異的衡量)。一旦建立了確定所有可能狀態之間相似性的標準,特徵數據就很容易轉換成距離數據。
對進化樹進行評估,主要採用Bootstraping法。進化樹的構
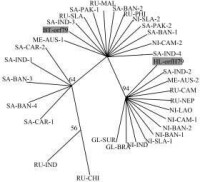
DNA系統發生樹
鄰接法是一個經常被使用的演演算法,它構建的進化樹相對準確,而且計算快捷。其缺點是序列上的所有位點都被同等對待,而且,所分析的序列的進化距離不能太大。另外,需要特別指出的是對於一些特定多序列對象來說可能沒有任何一個現存演演算法非常適合它。
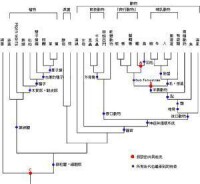
生物元素系統發生樹
基本信息
- 中文名
- 系統發生樹
- 外文名
- Phylogenetic tree
- 別名
- 演化樹
- 屬於
- 親緣分支分類方法