視桿細胞
位於眼球視網膜內的細胞
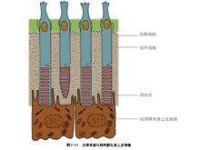
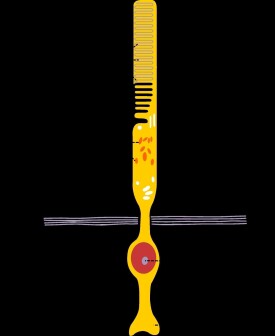
人類每個眼球的視網膜內約有1.2億個視桿細胞,其樹突呈細桿抓哏內,稱為視桿,視桿外節的膜盤除基部少數膜盤仍與胞膜相連,其餘大部分均在邊緣處與胞膜脫離,成為獨立的膜盤。膜盤的更新是由外節基部不斷產生,其頂端不斷被色素上皮細胞所吞噬。膜盤上鑲嵌有感光物質,稱視紫紅質(rhodopsin),能感受弱光。視桿細胞體較小,核圓形染色較深,其軸突末梢不分之呈球型,與雙極細胞的樹突形成突觸。
視桿細胞
,稱為視桿,視桿外節的膜盤除基部少數膜盤仍與胞膜相連,其餘大部分均在邊緣處與胞膜脫離,成為獨立的膜盤。膜盤的更新是由外節基部不斷產生,其頂端不斷被色素上皮細胞所吞噬。膜盤上鑲嵌有感光物質,稱視紫紅質(rhodopsin),能感受弱光。視紫紅質是由11-順視黃醛(11-cisretinae)和視蛋白(oposin)組成,前者是維生素A的衍生物,當維生素A缺乏時,視紫紅質合成不足,則患夜盲症。視桿細胞體較小,核圓形染色較深,其軸突末梢不分之呈球型,與雙極細胞的樹突形成突觸。
約翰·霍普金斯大學科學家領導的研究小組發現,眼睛感光的任務極有可能僅由三種細胞負責。
在2003年6月15日期《自然》雜誌的網路版上,研究小組報道,視錐細胞、視桿細胞和產生黑視蛋白的特殊視網膜細胞一起合作,包攬了小鼠對光強做出反應的全部工作。有其他研究人員提出產生感光色素cryptochrome的細胞也有感光作用,但霍普金斯的科學家表示,就小鼠而言,cryptochrome細胞沒有這個作用,在人類中恐怕也是如此。
“我們相當確信,視錐/視桿細胞系統和黑視蛋白系統是哺乳動物眼部唯一兩個感光系統。”約翰·霍普金斯大學基礎生物醫學科學研究所的神經科學教授King-Wai Yau博士說。“永遠不要說永遠,但至少目前還沒有證據表明有第三個系統存在。”
視紫紅質分解
在實驗中,Yau和博士后研究人員Samer Hattar博士除去編碼三種關鍵蛋白的基因,三種蛋白中的每一種都有助於傳遞來自視桿細胞、視錐細胞和黑視蛋白細胞的光信息。早先,阻止小鼠模型視桿細胞、視錐細胞和產生視力的細胞的光信息傳遞的傳統方法是使視桿細胞和視錐細胞退化。
“但我們想避免傳統的視桿、視錐細胞退化孝小鼠模型所帶來的不確定性。”Hattar說。“在那個模型中,你無法確定視桿細胞和視錐細胞喪失的百分比,也無法確定退化的視網膜本身是否會影響你的觀察結果。”
令分別除去了3種關鍵蛋白的單基因敲除小鼠進行雜交,最終產生的後代,也就是3基因敲除小鼠,每種蛋白的“劑量“都減半或者三種蛋白水平出現不同組合。
與倫敦帝國學院的Robert Lucas合作,霍普金斯大學的研究人員證明,3基因敲除小鼠根本無法調節瞳孔對光做出反應。在多倫多大學進行的實驗中,文章合著者Nicholas Mrosovsky發現,3基因敲除小鼠暴露於光照中時,活躍水平的調節也不適當。而三種蛋白各含一半劑量的小鼠在兩個實驗中反應都正常。
“即使其內部生物鐘告訴它現在是夜晚,正常小鼠暴露於強光時也會表現得不太活躍,會藏起來甚至睡覺。”Yau說。“這被看作是動機發生改變,也許小鼠意識到在光照中--即使生物鐘告訴它們現在是夜晚,它們被捕食的機會更大。但3基因敲除小鼠好像在白天與夜晚一樣活躍。”
研究人員設計了另外一個實驗來排除cryptochrome蛋白有感光作用,據報道該蛋白與果蠅的感光有關。由於每個光敏蛋白都是對光的特徵波長最敏感,因而將小鼠暴露於單波長光中可以揭示對眼睛的非視覺功能最重要的蛋白。
在這些在倫敦帝國學院進行的實驗中,研究人員發現傳統的視桿/視錐細胞退化小鼠當暴露於黑視蛋白細胞而不是cryptochrome細胞感應的光時,“重設”內部生物鐘的能力最強。
“在果蠅中,有兩個以上系統負責感光,而在斑馬魚和鳥類等一些動物中,感光甚至不只局限於眼睛,而這對於傳統的視桿/視錐細胞退化小鼠而言是不可能的事情。”Hattar說。Hattar正在區分視桿細胞和視錐細胞在感光中所扮演的角色。“但在哺乳動物中,所有這些功能--視力、生物鐘和光誘導活動,都存在於一個部位,那就是視網膜,並且僅由兩個系統、三類細胞完成。”
盲人的視桿細胞和視錐細胞喪失功能,但眼睛中產生黑視蛋白的細胞似乎仍能提供足夠的信息使身體和大腦與生物鐘協調一致。但如果完全失去雙眼,生物鐘就會遭到破壞。
視網膜神經細胞
哺乳動物光感受器細胞模式圖光感受器按其形狀可分為兩大類,即視桿細胞和視錐細胞。夜間活動的動物(如鼠)視網膜的光感受器以視桿細胞為主,而晝間活動的動物(如雞、松鼠等)則以視錐細胞為主。但大多數脊椎動物(包括人)則兩者兼而有之。視桿細胞在光線較暗時活動,有較高的光敏度,但不能作精細的空間分辨,且不參與色覺。在較明亮的環境中以視錐細胞為主,它能提供色覺以及精細視覺。這是視覺二元理論的核心。在人的視網膜中,視錐細胞約有600~800萬個,視桿細胞總數達1億以上。它們似以鑲嵌的形式分佈在視網膜中;其分佈是不均勻的,在視網膜黃斑部位的中央凹區,幾乎只有視錐細胞。這一區域有很高的空間分辨能力(視銳度,也叫視力)。它還有良好的色覺,對於視覺最為重要。中央凹以外區域,兩種細胞兼有,離中央凹越遠視桿細胞越多,視錐細胞則越少。在視神經離開視網膜的部位(乳頭),由於沒有任何光感受器,便形成盲點。
視網膜內有感光細胞層,人類和大多數脊椎動物的感光細胞有視桿細胞和視錐細胞兩種。感光細胞可通過終足和雙極細胞發生突觸聯繫,雙極細胞再和神經節細胞聯繫,由節細胞發生的突起在視網膜表面聚合成束,然後穿過脈絡膜和鞏膜后構成視神經,視神經出眼球后穿視神經管入顱腔,經視交叉連於間腦。
目前認為,物像落在視網膜上首先引起光化學反應,已從視網膜上提取出感光物質。這些物質在暗處呈紫紅色,受到光照時則迅速退色而轉變為白色。如將蛙或兔放在暗室中,使動物跟朝向明亮的窗子一定時間,然後遮光立即摘出眼球,剔出視網膜,用適當化學物質如明礬處理視網膜,則可發現動物視網膜留有窗子的圖像,窗子的透光部分呈白色,窗框部分呈暗紅色。這些都說明視網膜上感光物質在光線作用下所出現的光化學反應。在感光細胞的大量研究中,對視桿細胞研究得比較清楚。視桿細胞的感光物質稱為視紫紅質,它由視蛋白和視黃醛結合而成。視黃醛由維生素A轉變而來。視紫紅質在光照時迅速分解為視蛋白和視黃醛,與此同時,可看到視桿細胞出現感受器電位,再引起其他視網膜細胞的活動。
視紫紅質在亮處分解,在暗處又可重新合成。人在暗處視物時,實際上既有視紫紅質的分解,又有它的合成。光線愈暗,合成過程愈超過分解過程,這是人在暗處能不斷看到物質的基礎。相反在強光作用下,視紫紅質分解增強,合成減少,視網膜中視紫紅質大為減少,因而對弱光的敏感度降低。故視桿細胞對弱光敏感,與黃昏暗視覺有關。視紫紅質在分解和再合成過程中,有一部分視黃醛將被消耗,主要靠血液中的維生素A補充。如維生素A缺乏,則將影響人在暗處的視力稱為夜盲症。
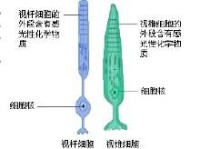
視桿細胞和視錐細胞
在外段小盤上排列著對光敏感的色素分子,這種色素通稱視色素,它在光照射下發生的一系列光化學變化是整個視覺過程的起始點。
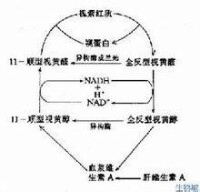
視桿細胞的視色素叫做視紫紅質,它具有一定的光譜吸收特性,在暗中呈粉紅色,每個視桿細胞外段包含109個視紫紅質分子,視紫紅質是一種色蛋白,由兩部分組成。其一是視蛋白,有348個氨基酸,分子量約為38000;另一部分為生色基團——視黃醛,是維生素A的醛類,因為存在若干碳的雙鍵,它具有幾種不同的空間構型。在暗處呈扭曲形的11-型異構體,但受光照后即轉變為直線形的全-反型異構體。後者不再能和視蛋白相結合,經過一系列不穩定的中間產物后,視黃醛與視蛋白相分離。在這一過程中,視色素分子失去其顏色(漂白)。暗處它在酶的作用下,視黃醛又變為11-順型,並重新與視蛋白相結合(復生),完成視覺循環。在強光照射后,視紫紅質大部分被漂白,其重新合成需要約1小時。隨著視紫紅質的復生,視網膜的對光敏感度逐漸恢復,這是暗適應的光化學基礎。當動物缺乏維生素A時,視覺循環受阻,會導致夜盲。
視紫質化學反應圖解
視桿細胞的信號和視錐細胞的信號,在視網膜中的傳遞通路是相對獨立的,直到神經節細胞才匯合起來。接收視桿細胞信號的雙極細胞只有一類(桿雙極細胞),但接收視錐細胞信號的雙極細胞,按其突觸的特徵可分為陷入型和扁平型兩種,這兩種細胞具有不同的功能特性。在外網狀層,水平細胞在廣闊的範圍內從光感受器接收信號,並在突觸處與雙極細胞發生相互作用。此外,水平細胞還以向光感受器反饋的形式調製信號。在內網狀層雙極細胞的信號傳向神經節細胞,而無長突細胞則把鄰近的雙極細胞聯繫起來。視桿和視錐細胞信號的匯合也可能發生在無長突細胞。
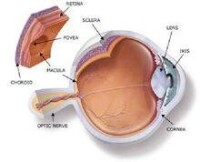
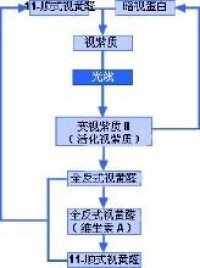
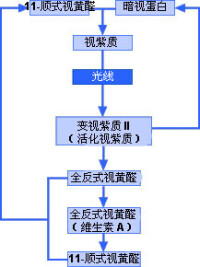
光線進入眼睛后,首先通過角膜,然後分別透過房水、晶狀體和玻璃體。最後,光線到達眼睛的感光組織視網膜。視網膜中有兩種細胞分別叫做視桿細胞和視錐細胞。視桿細胞負責昏暗光線下的視物,而視錐細胞則負責處理色彩和細節。當光線接觸到這兩種細胞時,會發生一系列複雜的化學反應。形成的化學物質(活化視紫質)會在視神經中產生電子脈衝。一般來說,視桿細胞的外段細長,而視錐細胞的外段更像是錐形。
這兩種細胞的外段都含有感光性化學物質。在視桿細胞中,這種化學物質叫做視紫質;而視錐細胞中的則叫做色素。光線進入眼睛后,會接觸到感光性化學物質視紫質(也叫做視紫紅質)。視紫質是暗視蛋白和11-順式視黃醛的混合物,11-順式視黃醛來源於維生素A。因此,缺乏維生素A就會導致視力問題。視紫質遇光會發生分解,因為光會導致視紫質中的11-順式視黃醛發生物理變化,轉化為全反式視黃醛。第一個反應只需一萬億分之幾秒的時間。11-順式視黃醛是角狀分子,而全反式視黃醛是直線分子。因此,化學性質極不穩定。視紫質可分解為幾種中間化合物,但最終(在不到一秒的時間裡)會形成變視紫質II(活化視紫質)。這種化學物質能夠產生電子脈衝,電子脈衝傳輸到大腦後,大腦將其解譯為光線。
視桿細胞
1.視桿細胞的細胞膜(外層)帶有電荷。當光線激活視紫質時,它就會導致環式GMP(鳥苷單磷酸)減少,而電荷隨之增加。這樣就會在細胞周圍產生電流。隨著光線的增多,就會有更多的視紫質被激活,也就會產生更強的電流。
2.最後,電子脈衝被傳到神經節細胞,再傳到視神經。
3.這些視神經在視交叉處彙集。在這裡,視網膜內部的神經纖維會通往另一側大腦,而視網膜外部的神經纖維則通往同側大腦。
4.這些纖維最終到達大腦後部(枕骨腦葉)。這個位置叫做基本視覺皮層,大腦在這裡將電子脈衝解譯為視覺信號。有些視覺纖維通往大腦的其他部分,有助於控制眼睛的活動、瞳孔和虹膜的反應,以及行為方式。
最後,必須重新形成視紫質,才能重複進行產生視覺的過程。全反式視黃醛轉化成11-順式視黃醛,然後與暗視蛋白結合后,便形成視紫質,視紫質遇光后便會重新開始該過程。
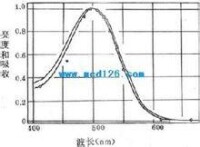
弱光條件下人眼所感到的光譜亮度曲線
(一)視紫紅質的光化學反應及其代謝
弱光條件下人眼所感到的光譜亮度曲線視紫紅質的分子量約為27-28kd,是一種與結合蛋白質,由一分子稱為視蛋白(opsin)的蛋白質和一分子稱為視黃醛(retnal)的生色基團所組成。視蛋白的肽鏈序列已搞清,它的肽鏈中有7段穿越所在膜結構、主要由疏水性氨基酸組成的α-螺旋區段,同一般的細胞膜受體具有類似的結構。視黃醛由維生素A變來,後者是一種不飽和醇,在體內一種酶的作用下可氧化成視黃醛。提純的視紫紅質在溶液中對500nm波長的光線吸收能力最強,這與人眼在弱光條件下對光就業上藍綠光區域(相當於500nm波長附近)感覺最明亮(不是感到了藍綠色)的事實相一致,說明人在暗視覺與視桿細胞中所含視紫紅質的光化學反應有直接的關係。
視紫紅質在光照時迅速分解為視蛋白和視黃醛,這是一個多階段的反應。目前認為,分解的出現首先是由於視黃醛分子在光照時發生了分子構象的改變,即它在視紫紅質分子中本來呈11-順型(一種較為彎曲的構象),但在光照時變為全反型(一種較為直的分子構象)。視黃醛分子構象的這種改變,將導致視蛋白分子構象也發生改變,經過較複雜的信號傳遞系統的活動,誘發視桿細胞出現感受器電位。據計算,一個光量子被視紫紅質吸收,就足以使視黃醛分子結構發生改變,導致視紫紅質最後分解為視蛋白和視黃醛。視紫紅質分解的某些階段伴有能量的釋放,但這看來不是誘發感受器電位的直接原因。
在亮處分解的視紫紅質,在暗處又可重新合成,亦即它是一個可逆反應,其反應的平衡點決定於光照的強度。視紫紅質再合成的第一步,是全反型的視黃醛變為11-順型的視黃醛,很快再同視蛋白結合。此外,貯存在視網膜的色素細胞層中的維生素A也是全反型的,它們也可在耗能的情況下變成11-順型的,進入視桿細胞,然後再氧化成11-順型的視黃醛,參與視紫紅質的合成補充;但這個過程進行的速度較慢,不是促進視紫紅制裁再合成的即時因素。人在暗處視物時,實際是既有視紫紅質的分解,又有它的合成,這是人在暗光處能不斷視物的基礎;光線愈暗,全盛過程愈超過分解過程,視網膜中處於合成狀態的視紫紅質數量也愈高,這也使視網膜對弱光愈敏感;相反,人在亮光處時,視紫紅質的分解增強,合成過程甚弱,這就使視網膜中有較多的視紫紅質處於分解狀態,使視桿細胞幾乎失去了感受光刺激的能力;事實上,人的視覺在亮光處是靠另一種對光刺激較不敏感的感光系統即視錐來完成的,后一系統在弱光時不足以被刺激,而在強光系統下視桿細胞中的視紫紅質較多地處於分解狀態時,視錐系統就代之而成為強光刺激的感受系統。在視紫紅質和再合成的過程中,有一部分視黃醛被消耗,這最終要靠由食物進入血液循環(相當部分貯存於肝)中的維生素A來補充。長期攝入維生素A不足,將會影響人在暗光處的視力,引起夜盲症。
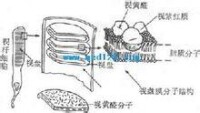
視桿細胞外段的超微結構示意圖
視桿細胞外段的超微結構示意圖感光細胞的外段是進行光-電轉換的關鍵部位。視桿細胞外段具有特殊的超微結構。在外段部分,膜內的細胞漿甚少,絕大部分為一些整齊的重疊成層的圓盤狀結構所佔據,這圓盤稱為視盤。每一個視盤是一個扁平的囊狀物,囊膜的結構和細胞膜類似,具有一般的脂質雙分子層結構,但其中鑲嵌著的蛋白質絕大部分是視紫紅質,亦即視桿細胞所含的視紫紅質實際上幾乎全部集中在視盤膜中。視盤的數目在不同動物的視桿細胞中相差很大,人的每個視桿細胞外段中它們的數目近千;每一個視盤所含的視紫紅質分子約有100萬個。這樣的結構顯然有利於使進入視網膜的光量子有更大的機會在外段中碰到視紫紅質分子。
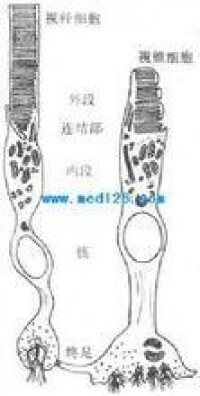
哺乳動物光感受器細胞模式
光子的吸收引起外段膜出現超極化電反應的機制已基本搞清,這就是光量子被作為受體的視紫紅質吸收后引起視蛋白分子的變構,又激海參了視盤膜中一種稱為傳遞蛋白(transducin)Ct的中介物,後者在結構上屬於G-蛋白家庭的一員,它激活的結果是進而激活附近的磷酸二酯酶,於是使外段部分胞漿中的cGMP大量分解,而胞漿中cGMP的分解,就使未受光刺激時結合於外段膜的cGMP由也膜解離而被分解,而cGMP在膜上的存在正是這膜中存在的化學門控式Na+通道開放的條件,膜上cGMP減少,Na+通道開放減少,於是光照的結果出現了記錄到的超極化型感受器電位。據估計,一個視紫紅質被激活時,可使約500個傳遞蛋白被激活;雖然傳遞蛋白激活磷酸二酯酶是1對1的,但一個激活了的磷酸二酯酶在一秒鐘內大約可使4千多個cGMP分子降解。由於酶系統的這種生物放大作用,就可以說明1個光量子的作用何以能在外段膜上引起大量化學門控式Na+通道的關閉,引起一個足以為人的視覺系統所感知的超極化型電變化。
視桿細胞外段和整個視桿細胞都沒有產生動作電位的能力,由光刺激在外段膜上引起的感受器電位只能以電緊張性的擴布到達它的終足部分,影響終點(相當於軸突末梢)外的遞質釋放。
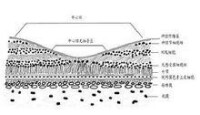
感光色素
產生視覺形象是眼睛最為人熟知的工作,但感光並對光水平做出反應也是眼睛一項極為重要的工作。有了這個附屬能力,眼睛就可以維持機體內部生物鐘運轉,使瞳孔對光有反應,在一天中適當的時間激活響應的活動動機。科學家報道,沒有這三種細胞發送的信號,小鼠就會喪失這些正常能力。
在實驗中,Yau和博士后研究人員Samer Hattar博士除去編碼三種關鍵蛋白的基因,三種蛋白中的每一種都有助於傳遞來自視桿細胞、視錐細胞和黑視蛋白細胞的光信息。早先,阻止小鼠模型視桿細胞、視錐細胞和產生視力的細胞的光信息傳遞的傳統方法是使視桿細胞和視錐細胞退化。
“但我們想避免傳統的視桿、視錐細胞退化孝小鼠模型所帶來的不確定性。”Hattar說。“在那個模型中,你無法確定視桿細胞和視錐細胞喪失的百分比,也無法確定退化的視網膜本身是否會影響你的觀察結果。”
研究人員設計了另外一個實驗來排除cryptochrome蛋白有感光作用,據報道該蛋白與果蠅的感光有關。由於每個光敏蛋白都是對光的特徵波長最敏感,因而將小鼠暴露於單波長光中可以揭示對眼睛的非視覺功能最重要的蛋白。
在這些在倫敦帝國學院進行的實驗中,研究人員發現傳統的視桿/視錐細胞退化小鼠當暴露於黑視蛋白細胞而不是cryptochrome細胞感應的光時,“重設”內部生物鐘的能力最強。
基本信息
- 中文名
- 視桿細胞
- 外文名
- rod cell
- 類型
- 視覺細胞
- 作用
- 分辨黑白,動態信息