光合色素
光合色素
光合色素(photosynthetic pigment) 在光合作用中參與吸收、傳遞光能或引起原初光化學反應的色素。光合色素存在於葉綠體類囊體膜,包含葉綠素、反應中心色素和輔助色素。
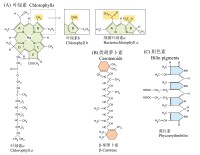
一些光合色素的分子結構
高等植物和大部分藻類的光合色素是葉綠素a,b和類胡蘿蔔素;在許多藻類中除葉綠素a,b外,還有葉綠素c,d和藻膽素,如藻紅素和藻藍素;在光合細菌中是細菌葉綠素等;在嗜鹽菌中則是一種類似視紫質的色素11-順-視黃醛(11-cis-retinal)。

光合色素
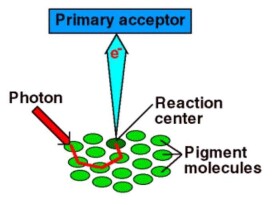
依功能不同,光合色素可分成天線色素和反應中心色素兩類。天線色素捕獲光能,並將光能傳給反應中心。極大部分光合色素都起這一作用。反應中心色素的作用是以光能來引起電荷分離及光化學反應。它的主要成分是特殊的葉綠素a,其存在狀態和光譜性質不同於一般的葉綠素a。光合色素所以能表現其特殊功能,是由於它在光合器中以特定的形式和蛋白質、脂質等結合。結合態的光合色素的性質如吸收光譜、氧化還原電位等,和非結合態的有明顯差別。例如,葉綠體中的葉綠素a的紅光波段吸收峰與在丙酮溶液中時相比,向長波方向偏數十納米。光系統Ⅰ(PSⅠ)的葉綠素在照光時,在700納米處有光吸收變化,光系統Ⅱ(PSⅡ)的則在680納米處有光吸收變化(因而把光系統Ⅰ,Ⅱ分別稱為P700和P680)。在PSⅠ中的葉綠素 a的氧化還原電位比離體時測得的數值低得多。
葉綠素chlorophyll 存在於光合作用生物類囊體中的同化色素的一種。在高等植物C3植物的葉綠體中,葉綠素a、b兩類大致以3∶1存在。在中央具有一個Mg原子的二氫卟吩(dihydroporphin)的衍生物上,葉綠素a、b是甲醇及葉綠醇與酯結合的物質。葉綠素a及b用稀酸處理, Mg被H置換,而變成去鎂葉綠素a、 b。另外通過葉綠素酶脫掉葉綠素分子中的植基(Phytyl),就產生脫植基葉綠素a、b(chlorophyllide)。通常在黃化葉的原生質體中有原脫植基葉綠素(proto-chlorophyllide)代替葉綠素,通過光照射,它又變成為脫植基葉綠素,最後形成葉綠素。現已知除光合細菌外,所有光合生物都含有葉綠素a。高等植物和紅藻含有葉綠素b,硅藻和褐藻含有葉綠素c,紅藻的某些種含有葉綠素d。光合細菌中含有細菌葉綠素a、b、c、d。葉綠素a的固體是黑色,其溶液是藍綠色。葉綠素b的固體是綠黑色,溶液是綠色。葉綠素在紅色與藍紫色部分表現出明顯的吸收光譜。葉綠素a、b、c、d在乙醚溶液中的紅色與藍紫色部分吸收最大(毫微米),分別為662,430;644,455; 628,447; 688,447(參見圖)。在生活細胞中,它們的主要吸收帶和在溶媒中比較,是在10—50毫微米波長範圍內變動。根據活細胞的分光光度學的研究,已知在類囊體中至少有四種形態的葉綠素a存在。葉綠素在有機溶劑中發生強烈的熒光。在用光照射活細胞時所見到的葉綠素熒光,遠比在溶媒中的少。
葉綠素是植物中進行光合作用的主要色素,是一類含脂的色素家族,位於類囊體膜,並且賦予植物的綠色。葉綠素吸收的光主要是藍紫色和紅色而不是綠色光,(綠光吸收量少,故葉綠素呈綠色)它在光合作用的光吸收中起核心作用。葉綠素分子是由兩部分組成的:核心部分是一個卟啉環(porphyrin ring),其功能是光吸收; 另一部分是一個很長的脂肪烴側鏈,稱為葉綠醇(phytol),葉綠素用這種側鏈插入到類囊體膜。與含鐵的血紅素基團不同的是,葉綠素卟啉環中含有一個鎂原子。葉綠素分子通過卟啉環中單鍵和雙鍵的改變來吸收可見光。
植物含有幾種類型的葉綠素,它們之間的差別在於烴側鏈的不同。葉綠素a存在於能進行光合作用的真核生物和藍細菌中。大多數能進行光合作用的細胞還有第二種類型的葉綠素,即葉綠素b 或葉綠素c。在高等植物和綠藻的細胞中含有葉綠素b,而在其它一些類型的細胞中含有葉綠素c。不同類型的葉綠素對光的吸收也是不同的,如葉綠素a最大的吸收光的波長在420~663nm,葉綠素b 的最大吸收波長範圍在460~645nm。當葉綠素分子位於葉綠體膜上時,由於葉綠素與膜蛋白的相互作用,會使光吸收的特性稍有改變。
類胡蘿蔔素(carotenoid)是一類由8個異戊二烯單位組成的,含有40個碳原子的化合物,不溶於水而溶於有機溶劑。葉綠體中的類胡蘿蔔素含有兩種色素,即胡蘿蔔素(carotenoid)和葉黃素(xanthophyll),前者呈橙黃色,後者呈黃色。一般情況下,葉片中葉綠素與類胡蘿蔔素的比值約為3:1,所以正常的葉子呈現綠色。而在葉子衰老過程中,葉綠素較易降解,而類胡蘿蔔素比較穩定,所以葉片呈現黃色。
藻膽素(phycobilin)是藻類主要的光合色素,僅存在於紅藻和藍藻中。常與蛋白質結合為藻膽蛋白。藻膽素的4個吡咯環形成直鏈共軛體系,不含鎂和葉綠醇鏈,具有收集和傳遞光能的作用。
基本信息
- 中文名
- 光合色素
- 外文名
- photosynthetic pigment
- 存在於
- 葉綠體基粒
- 韓語
- 포토 크롬
- 西班牙語
- Pigmentos
- 法語
- Pigments photosynthétiques