蛻皮動物
蛻皮動物
蛻皮動物(Ecdysozoa)是一大類原口動物,包括節肢動物門、線蟲動物門和幾個徠小門,最初由Auinaldo等人於1997年定義,主要根據是18S核糖體RNA樹。而這個分類同時也被一系列形態學證據所支持,因為這個分類包括所有蛻掉外骨骼的動物(見蛻皮(ecdysis))。與蛻皮動物接近的定義也曾經被Perrier在1897年和Seurat在1920年僅依據形態作出。

蛻皮動物
蛻皮動物包括如下門:節肢動物門、有爪動物門、緩步動物門、動吻動物門、鰓曳動物門、鎧甲動物門、線蟲動物門和線形動物門;其它一些門類,比如腹毛動物門,曾被認為可能是其中的一員,但因其缺乏蛻皮動物的主要特徵,現在通常被劃出。泛節肢動物的共同特徵是具有分節的身體,原來以為由環節動物演變而來,而與其一起組成分節動物(Articulata)。然而它們並不具有太多的共同特徵,現在看來他們分別演化出了分節的特點。此外,從18S 核糖體RNA樹來看,后口動物中的毛顎動物門可能與其餘后口動物不同而屬於蛻皮動物。蛻皮動物中不屬泛節肢動物的成員曾經被組成 環神經動物(Cycloneuralia),但通常被認為是並系群。
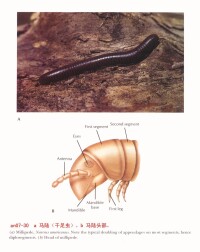
蛻皮動物-節肢動物門
葉足動物門(Lobopodia):奇蝦 歐巴賓海蠍 微網蟲 埃謝櫛蠶 怪誕蟲等等。
節肢動物門(Arthropoda):蜘蛛,蜈蚣,蟬等。
有爪動物門(Onychophora):毛蟲等。
緩步動物門(Tardigrada):水熊蟲,緩步蟲等。
動吻動物門(Kinorhyncha)

蛻皮動物
鎧甲動物門(Loricifera)
線蟲動物門(Nematoda):鉤吻球頸線蟲,人蛔蟲等等。
蛻皮動物是指所有會產生蛻皮現象動物的總稱,其中還包括線蟲、節肢動物和蠕蟲( Aguinaldo et al.1997),兩個被研究得最為透徹的的分子生物學模式生物線蟲和果蠅都屬於蛻皮動物,在強大的遺傳學研究技術支持下,這兩種模式生物為生物學研究做出了巨大貢獻,其中涵蓋了從細胞基本原理到系統學分析等一系列主題,而且,這些研究對我們理解生物學的重要性毋庸置疑。雖然上面羅列了這些研究的意義,但越來越多的證據正在挑戰以下這些不言而喻的假說:這些研究結果能適用於大部分的無脊椎動物,或可以通過這些研究結果來推斷較簡單的祖先動物的代謝過程。相反,這些數據表明,現存的模式蛻皮動物的一些簡單特徵是祖先複雜結構發生二次簡化的結果。
儘管一些蛻皮動物類群很少保留祖先的複雜性,但它們仍進化出迷人的複雜特徵。例如,果蠅唐氏綜合征細胞粘附分子(Dscam)基因的內含子含量很高,在所有已鑒定的生物中剪接變異體數目最多(Schmucker et al.2000)。黑腹果蠅的Dscam能將特定的物質傳給遷移神經元,並協調同種抗原排斥,這是建立神經迴路的重要前提。Dscam基因的轉錄產物在一些群體(包括甲殼動物水蚤)中表現出較高的多樣性,表明Dscam基因可變剪接導致細胞多樣性的形成,這可能是節肢動物擁有的一個基本功能(Brites et al.2008)。與此相反,脊椎動物中Dscam同源基因似乎並沒有出現廣泛的可變剪接,雖然它們似乎也存在有同種抗原相互作用現象(見綜述Hattori et al.2008)。
這個例子說明,在衡量動物的複雜性時,即使是基於定量分子分析,仍需要區分它們是保留了祖先的複雜特徵還是二次得到的複雜特徵。還不清楚這兩種變化方式的相互關係,因此判定某個物種或類群的複雜性時需要估計它們的進化起源時間。同樣值得注意的是,昆蟲和線蟲都具有非常高的多樣性,而且昆蟲是動物中物種最豐富的類群(Brusca and Brusca 2003),儘管昆蟲丟失了一些複雜的特徵,它們卻進化出了非常高的形態複雜性和多樣性。但是,在海洋環境中昆蟲的多樣性並不算很高,其他蛻皮動物類群(包括甲殼類動物和自由生活的線蟲)在海洋中丰度更高,然而適用於它們的悔洋模型和分子學數據仍然非常稀缺。
