大腦半球
大腦半球
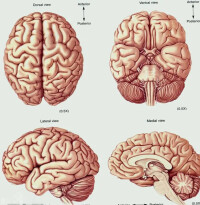
大腦半球(cerebral hepIMC),端腦的兩側結構。兩半球前後完全分開,中間由一束最大的連合纖維——胼胝體連接。表面是灰質,即大腦皮層,其隆起的部分為腦回,凹陷的部分稱溝裂。背外側面是由大腦外側裂和中央溝等分成的額、頂、枕、顳等腦葉。最前端是額極,最後端是枕極。皮質下是厚的白質,即大腦髓質。據構成的路徑及其聯繫,髓質纖維柬可分為三類:投射纖維、半球內的聯合纖維和半球間的連合纖維。投射纖維是皮質與皮質下中樞的聯絡纖維,包括傳導衝動到皮質的上行纖維束以及將衝動從皮質傳到皮質下中樞的下行纖維束。上行纖維主要是丘腦放射、昕放射和視放射,下行纖維主要為錐體系和錐體外系。
大腦半球
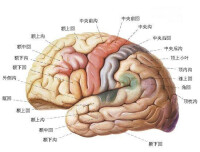
在半球表面有各種走向的溝即大腦溝(sulcus cerebri),由其主要的溝或裂把每一半球分為額葉(lobus frontalis)、頂葉(lobus parietalis)、枕葉(lobus occipitalis)、顳葉(lobus temporalis)和腦島(insula)。腦島被其他葉所遮蓋,在自然位置條件下看不見。額葉的外側面進一步由數條溝將之分為中央前回(gyrus praecentralis)。額上回(gyrus frontalis superi-or)、額中回(gyrus frontalis medius)、額下回(gyrus frontalis inferior),下面位於眼窩的近上方,在內側有前後走行的嗅溝,該處有嗅球(bulbus olfactorius)和嗅束(tractus olfacto-rius),為由嗅覺神經末稍向中樞的通路。頂葉的外側面與中央溝(sulcus centralis)為界,在額葉的後方分為中央后回(gyrus postcentralis)、頂上葉(lobulus parietalis superior)、頂下葉(lobulus parietalis inferior),但後者又分為前端的環曲回(gyrus circumflexus)和後端的角回(gyrus angularis)。枕葉的溝和回都不規則,但猿類所存在的猿回(德 Affenspalte)在人類有的也有。顳葉是頂葉後下部和枕葉的向前外側突出的部分,在額葉和頂葉前部之間有大腦外側裂(fissura cerebri lateralis)。
外側面可分為顳上回(gyrus temporalis superior)、顳中回(gyrus temporalis medius)和顳下回(gyrustemporalis inferior),而顳上回的內側則稱為顳橫回(gyri temporales transversi),為聽覺中樞所在的部位。構成顳葉內側面一部分的海馬回溝(un-cus gyri hippocampi)或海馬溝(sulcus hippo-campi)相當於原皮質,為嗅覺中樞的位置。在大腦半球的內側面有從上方包著胼胝體的扣帶回(gyru-cinguli),在後方尚有距狀溝(sulcus calcarinus)過枕葉內側面後行,在此附近構成視覺中樞。人的大腦半球質有100億以上的神經細胞,大致作一定的層次排列,由神經細胞發出的神經纖維構成白質而達至其他部位。
在這些纖維中,成為同一半球內從一皮質區到另一皮質區連繫通路的稱為連繫路,(德Assoziationsbahn)而把通過胼胝體向另一半球相同部位的連繫通路稱為連合路(德Kommissurenbahn),通過大腦核、內囊(capsulainterna)向大腦半球以外的通路為投射路。(德Proje-ktionsbahn)在左右大腦半球的內部,每側腦室的前部各形成側腦室(ventriculi laterales),充滿腦脊髓液。人的大腦核,可區分為:沿側腦室內面存在的尾狀核、腦島內側的豆狀核(nucleus lentifofmis由殼和蒼白球構成)、屏狀核(claustrum)和位於顳葉前端部的杏仁核。
大腦是人體的一個器官,它比世界上最高級的電腦還要複雜和充滿奧秘。人腦重約3磅,它由兩部分構成:左半腦和右半腦。這兩個部分通過胼胝體相連接。胼胝體實際上是一束神經組織,負責協調左、右半腦的工作。它使兩個半腦發生聯繫,使記憶和學習的傳輸活動得以實現。
人腦的兩個部分:左腦和右腦
看上去大腦的兩個部分——左半腦和右半腦長得很對稱,就像互相在照鏡子。然而,它們實際上是非對稱組織,換句話說,它們在結構和功能上有著諸多不同。
左腦和右腦的功能分工
運動原皮層分佈在左、右半腦的頂部,在結構上呈兩邊對稱。運動原皮層下面是感覺區,同樣有兩邊對稱的結構,負責接收和處理各種信息,這些信息來自皮膚、骨骼、關節、肌肉以及肢體的運動。這一區域有時也被稱為觸覺區。大腦的運動區和感覺區的分工,是很專門化的:每一個特定區域都負責控制身體某一特定部位,左半腦控制右手和右腳,而右半腦則控制身體的左側。
大腦皮層的軀體感覺區和運動區
大腦皮層的這兩個區域的每一個部位都與身體的某一部分發生聯繫。身體的大多數器官和組織都在大腦皮層的相應部位得到顯示。控制身體某一部分的大腦皮層相應區域的大小,與身體這一部分的實際大小無關,而與控制它所需的精密程度成正比。人腦用來控制臉和手的區域比較大,而控制肩、頸、臀、軀幹等的區域相比較小。
枕葉位於後腦緊挨腦幹的上方,是產生視覺的地方。儘管每個半腦都接收來自左、右兩個視野的信息,但左眼發送更多的信息給右半腦,右眼則發送更多的信息給左半腦。每一隻眼睛都有一個相對應的優勢半腦。所謂優勢半腦,是指對信息更多地作出反應或起支配反應的那個半腦。
大腦半球
如前所述,大腦並非完全對稱。許多專門的功能性中樞似乎主要在大腦的這一半球,或者另一半球。說話中樞和聽覺中樞在大腦左側緊挨耳朵上方的部位(Broca區)。聲音記憶區在聽覺中樞的後面。語言區(即Wernicke區)對大多數人來說是在左半腦,但其實它在兩個半腦中的任何一個都能形成。
大腦兩半球功能上的不對稱,或者說腦的不同功能向一側半球集中是人腦結構和認知的主要特徵,生理學上稱之為大腦半球一側優勢,或簡稱大腦優勢。在98%以上的成年右利手者中,左半球專管對語言的處理和語法表達,如詞語、句法、命名、閱讀、寫作、學習記憶等。而空間技巧與右半球相關,如對三維形狀的感知、空間定位、自身打扮能力、音樂欣賞及歌唱等。右半球還可理解一些口語及印刷的詞。可以認為左半球是科學性的,而右半球是藝術性的。大腦半球一側優勢在成人有,兒童有,嬰兒也有,甚至某些動物也有。
1863年,法國外科醫生皮埃爾·布羅卡(PierreBroca)指出兩個大腦半球的功能有差別,左額葉可能是控制言語的皮層區。而除了左半球言語功能佔優勢外,人們認為兩個半球在感覺和運動功能方面全是對等的。左半球接受身體右半側的感覺傳入,並支配右半側肌肉運動;右半球接受身體左半側的感覺傳入,並支配左半側肌肉運動。
20世紀50年代,美國加利福尼亞州技術研究院教授斯佩里真正確立了左右腦分工的觀念,他與學生嘗試把貓,猴子左右腦連接的神經線全部切斷後,這些動物仍然生活得正常,並且可以訓練兩個腦半球以相反的方式去完成同一項任務。研究結果顯示,左腦同樣具有右腦的功能,右腦也同樣具有左腦的功能,只是分工和側重點不同。
對大腦優勢的臨床研究,最先始於對裂腦人的實驗觀察。自然腦損傷可造成裂腦,在腦損傷病人身上觀察到,左側腦損傷導致右側腦功能喪失,主要是語言功能的喪失,但不影響右側腦功能;同樣,右側腦損傷導致左側腦功能喪失,患者可表現穿衣失用症,因分不清左右側而穿倒衣服,不能繪製圖表,視覺認識出現障礙,說明左右大腦半球是獨立活動的或功能分離的。裂腦人的主要來源是癲癇病人,為防止發病時左右兩半球間的傳播發作,減弱癲癇發病強度,常採取切斷病人胼胝體的方法,術后病人便成為裂腦人。將圖片在裂腦人的左半視野閃過,病人不能說出圖片上物體的名稱,因為視覺形象投射到了右半球而右半球是不具有說話功能的,但病人可用一些非言語形式表明他們已感知到了物體,比如用手收集和圖片上一樣的物體。說明病人的右側視覺是良好的,更重要的是說明了語言中樞位於左半球。而在正常人,由於胼胝體的作用,使兩側半球功能得以聯繫,因此不論物體出現在哪側視野,都可用詞語說出物體的名稱。
以正常人為實驗對象,則主要通過對那些與左右側大腦認知神經生理學相關的正常行為進行研究來闡明大腦優勢的存在,如側面呈現刺激物的感知實驗,實驗結果提示,當腦的一側對某一認知活動參與越多時,其注意力偏向對側感受野的活動就越大,從而引起感覺不對稱的增加。最近還採用在特殊感知活動期間進行觀察的腦成象技術,如腦局部血流圖和正電子發射斷層掃描術等。這些技術可以指示認知期間神經活動增強的部位,但神經活動增強標誌著什麼至今仍不清楚。研究正常個體,對象易得,個體間的差異如性別、年齡、認知水平、家族史、手偏利等的研究可行,左右大腦差異定量打分使研究半球不對稱的程度成為可能。
結構上來說,人腦右半球略重於左半球,但左半球灰質多於右半球;左右顳葉有明顯不對稱性;顳葉的不對稱性和丘腦的左右不對稱性相關;各種神經遞質的分佈,左右半球也是不平衡的。
從功能上來說,在正常情況下,大腦兩半球是協同活動的,進入大腦任何一側的信息會迅速通過胼胝體傳達到另一側,做出統一的反應。割裂腦研究表明大腦兩半球可能具有不同的功能。語言功能主要定位在左半球,負責語言、閱讀、書寫、數學運算、和邏輯推理等。而知覺物體的空間關係、情緒、欣賞音樂和藝術則定位於右半球。
大腦的功能存在單側優勢,但不是絕對分離的。近年來許多研究發現,右半球在語言理解中同樣也起重要作用。在加工複雜程度不同的句子時,右半球上與左半球經典語言區對應的部分也得到激活,只是激活的強度低於左半球。
本段總結自《普通心理學》(修訂版) 彭聃齡 2001
腦功能不對稱的理論:大腦一側優勢與遺傳有一定關係,但主要是在後天生活實踐中逐步形成的。在2~3歲時,左右側腦損傷的結果相差不多,因為那時尚未建立一側優勢;10~12歲時,如左半球損傷,可在右側半球建立語言中樞;在成年人,左側語言優勢已建立,左半球的損傷導致不可補償的語言障礙。另外,早期腦損傷的功能恢複比晚期腦損傷的功能恢復快,因為在生命早期神經系統的可塑性大,那時大腦半球還未特化或特化不明顯。
從解剖學圖形上看,大腦兩半球並不是鏡映的,這是功能不對稱的基礎。在第一顳回上的表面後部即顳面有著最可靠的形態學不對稱,而這兒正是語言綜合必需的核心部位。左顳面較大,超過右顳面約40%。其他不對稱的部位如左側大腦外側裂較長,仰角水平低,后額頂區較大,枕葉後端較寬等。左右半球功能不對稱的本質是什麼呢?一開始人們用言語-非言語或言語-視覺來區別左右半球的功能分離。這種分類不能解釋一些較新的資料,如在某些具有空間特徵的刺激中,對英文手寫體的識別與右半球的功能相關,而對印刷體和東方國家的表意文字的識別則與左半球相關。再如,感覺那些具有序列和時間順序的非言語刺激更依賴於左半球而不是右半球,而對某些聽刺激如音樂旋律的感覺更多地依賴於右半球。因此,兩半球不對稱的本質差異更準確地被描述為分解-合成或時間-圖形的功能分離。即左半球對不同條目根據時間順序安排的分解刺激加工的信息是特化的,如語言、語法技巧;右半球則對合成刺激加工的信息是特化的,並繼續形成統一的圖象,如對旋律、三維物體的感知。當然,左右半球的功能不對稱存在個體差異,比如在缺乏音樂素養的個體中,旋律感覺往往取決於右半球的作用,但在音樂家中則包括左半球的作用,反映出音樂家能較大地利用大腦的分析過程。
兩半球間的差異是絕對的或是相對的?即不清楚每個半球是否具有另一半球所特化的加工信息的能力。雖然兩半球都有處理語言刺激的能力,但並不意味著處理方式完全相同。左半球可能以分析模式進行加工,右半球則以整體模式進行加工。我們只能辯證地認為兩半球間的差異是相對的而不是絕對的,但缺乏充分的實驗證據。另外,我們也不清楚大腦功能不對稱究竟在神經系統的什麼水平存在。
動物腦是否具有功能不對稱性?按傳統概念,語言被認為是大腦功能不對稱的基礎,動物沒有明顯的語言,因此沒有腦功能不對稱。但近年研究表明,動物大腦存在功能不對稱,靈長類的左右半球間存在解剖學和功能上的不對稱。如黑猩猩的左側外側裂比右側裂要長,但二者差異比人類的要小。在發聲動物金絲雀中,左半球損傷對鳴叫的影響比右半球損傷大得多。除靈長類及鳥類外,其它動物如貓、兔、大鼠、小鼠都有腦功能不對稱效應。在猴及鼠還存在爪偏利,與人類手偏利的不同在於其右利及左利的個體數量基本相等,而在人類,右利手者明顯居多。這些觀察對以語言為大腦功能不對稱基礎的傳統理論提出了挑戰。可見,對腦功能不對稱的神經生物學基礎的闡明還僅僅是個開始。
中科院生物物理研究所腦與認知科學國家重點實驗室最新研究發現:人類大腦左半球對大範圍拓撲性質的知覺佔優勢,右半球對局部幾何性質的知覺佔優勢。該發現發表於2007年12月26日的《美國科學院院刊》,並被該刊《本期導讀》專欄介紹。
中科院生物物理所陳霖院士領導的認知科學和腦成像團隊,創立了“大範圍首先”的拓撲性質知覺理論,向半個世紀以來占統治地位的“局部首先”的理論提出挑戰,強調知覺過程是由大範圍拓撲不變性質開始的,為左右大腦差別的研究提供了一條嶄新的思路。
該所王波等青年學者把“大範圍首先”的拓撲性質知覺理論應用到大腦半球不對稱性的研究。他們經過6年多的大量實驗把各種拓撲性質(如洞的個數、內外關係等)和其他幾何性質(如朝向、距離、大小、對稱性、平行性、直線性等)進行了系統比較測試,一致揭示了慣用右手的人左腦拓撲性質的知覺佔優勢;還用功能磁共振成像發現,拓撲性質分辨產生大腦左半球顳葉興奮。有專家認為,該研究得出的結論“左半球拓撲性質知覺佔優勢、右半球局部幾何性質知覺佔優勢”,為解決視覺的左右大腦關係的各種爭論問題提供了一個統一的理論框架,同時為認識大腦、開發大腦提供了一個基本的科學根據。

基本信息
- 中文名
- 大腦半球
- 外文名
- pallium
- 釋義
- 大腦半球為腦的一部分
- 構成
- 內部的大腦核和外部的腦皮