古海洋學
古海洋學
古海洋學是深海鑽探計劃(DSDP)的產物,15年的(1968~1983年)鑽探不僅證實了板塊學說,而且產生了古海洋學這門新學科。古海洋學以大洋的水體作為主要研究對象,探索海洋環流和海上物理,化學特徵的變化。研究海洋生產力和海洋生物宏觀演化等歷史。現代含義上的古海洋學產生於70年代中期,當時深海鑽探的結果已經提供出古海洋學研究的典型成果。此後的近20年,古海洋學成為地球科學中發展最快的分支之一。

(圖)古海洋學
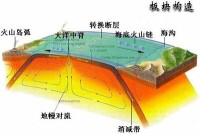
(圖)板塊構造
古海洋學作為一門獨立的學科出現是20世紀70年代以後的事情。1968年開始實施深海鑽探計劃,許多鑽井在各深海大洋區鑽穿整個沉積層達到基底。70年代末液壓活塞取心技術的出現,能獲取大量未擾動的長岩心,為研究中生代以來的海洋環境提供了較為精確的資料。大洋生物地層學,特別是微古生物及超微化石的研究進展迅速,它與磁性地層學、同位素年代學、岩性地層學以及火山灰年代學等技術結合起來,使得全球性地層時代對比和環境分析成為可能。板塊構造說為古海洋環境的再造提供了理論基礎。古生物學、地球化學及礦物學的現代科學技術廣泛應用於古海洋環境的研究,大大推進了古海洋學的發展。利用古海洋學理論和方法,初步建立了中生代以來各時期的海洋古環流、海洋古地理和古氣候演變的模式。
研究方法

(圖)微體古生物雜誌
微體古生物法 是古海洋學最主要的研究手段。有孔蟲、放射蟲、硅藻、顆石藻等微體或超微體生物的生活,主要受海水深度、溫度、鹽度、濁度、營養鹽以及水體運動等各種物理化學條件的控制。這些要素變化的信息便記錄在生物個體、生物組合、分異度等特徵上,因此海洋生物是海洋環境的靈敏標誌。微體古生物方法主要從以下幾方面進行分析:①生物時空分佈規律的研究。不同生物對其生活環境有一定選擇性,如放射蟲多見於赤道海域,硅藻多產於高緯度海區,窄溫性有孔蟲,有的適應於溫水,如截錐圓輻蟲(Globorotalia truncatulinoides);有的適應於冷水,如厚壁新方球蟲(Neogloboquadriua pachyderma)。根據生物分佈還可以推斷古海岸線的位置。從底棲有孔蟲的居住帶和生物分異度可推斷古水深。微體生物化石組合和延伸方向大體指明水團和海流流向。窄鹽性動植物化石可作為判斷海水鹽度的指標。生物分異度和生物組合與溫度梯度有關,分異度隨著緯度升高而降低。海洋動植物的緯向分帶標出了氣候帶。②生物個體形態特徵的研究。生物殼體的形態、大小、厚度、密度、旋轉方向及骨骼孔隙度等變化,都是為適應生活環境而發生的,它們主要反映水深及水溫的變化情況。生物殼體厚度的增減規律與靜水壓力有關,據此可判明古水深。浮游生物骨骼的孔隙度隨著水的密度增大而減小。一些浮遊有孔蟲殼的旋轉方向隨溫度發生變化,在冷水中多為左旋,暖水中多為右旋,可以用來判別季節變化和氣候帶。底棲有孔蟲的平均壽命在高緯度區比低緯度區大2~3倍。③植物光合作用嚴格受到海水深度的控制,而海水透光帶一般局限在表層200米內,故根據一些植物化石可以判斷古海水深度。
地球化學方法 利用海洋沉積物中某些元素和同位素的含量及其比值,可以確定古海水溫度、鹽度及水團。
同位素指標 由於海洋生物殼體的18O/16O比值,一方面隨殼體形成時的海水溫度而變化,另一方面隨當時海水同位素組成(與極地冰蓋的消長有關)而變化,因此通過海洋生物化石中氧同位素比值 (18O/16O)的測定,可以計算古海水溫度並推斷古氣候。目前,用於同位素測定的海洋生物主要是有孔蟲,此外還有顆石藻、軟體動物和珊瑚等化石。海水蒸發作用也能引起氧、碳同位素比值的變化。因此它們也可以用來估算海水鹽度。
化學元素指標 某些化學元素,特別是痕量及微量元素的溶解度和吸附量隨著海水溫度和鹽度發生變化,也可作為古海水溫度和鹽度的指標。如方解石中的Mg、Sr的富集隨溫度而增加,惰性氣體Ar、Kr、Xe的溶解度隨溫度的增加而減小。粘土礦物吸收硼的量與鹽度成比例,一般海洋沉積物比淡水沉積物含有更多的硼。海水中的許多痕量及微量元素丰度比淡水中的大,對這些元素進行統計分析可估算古鹽度。海洋碳酸鹽中的Na和Sr含量大於淡水的,海水的Sr/Ca和Fe/Mn值遠大於淡水的。海洋有機質 C/N比是4.3~7.1,反映了以海洋浮游生物(C/N為5~6)為主,而陸源植物的C/N則為30~40。因而它們都是判斷古鹽度的標誌。
沉積學方法 沉積物的礦物、化學成分、結構、構造特徵及其空間分佈,都可以用於判斷古海洋環境。
特徵礦物 文石或高鎂方解石多半出現於淺海環境;而低鍶或低鎂方解石出現於淡水環境。菱鐵礦多半產於10~50米水深的淺海區。自生黃鐵礦一般代表海水流動微弱或停滯的還原環境。呈帶狀分佈的石英、高嶺石、伊利石等陸源物質和火山灰往往被用來解釋海流狀況。
岩性特徵 碳酸鹽補償深度在地質時期歷經波動,根據它的波動情況,可以推斷古海水深度、生物生產力、水化學特徵以及海平面變動情況。蛋白石質二氧化硅、磷酸鹽的含量和分佈可指示某一時期上升流和生物生產力的狀況。深海地層中錳結核的出現,岩性突變、地層和化石帶的缺失等,可用來指示沉積間斷;而區域沉積間斷常可說明地質時期深海底層流及其侵蝕作用的存在。富含有機質的黑色頁岩,則可能代表水流微弱或停滯的缺氧還原環境。
粒度和構造特徵 利用沉積物粒度及其排列方向可以測定古海流的強度和流向,近年來有人試圖利用碎屑顆粒長軸導向磁化率來測定古海流方向。有時還可以利用粒度分配特徵,大體區分古氣候帶,例如,以機械風化為主的高緯度區多出現礫石等粗碎屑物質,常伴有分選很差的冰載物質;而以化學風化為主的低緯度帶則以泥質沉積為主。根據冰載物質在洋底沉積中的分佈狀況,尚可用來追索當時洋流的路徑與方向。地層中的濁流沉積物往往代表古海洋某種異常事件,如地震、風暴、滑坡等。
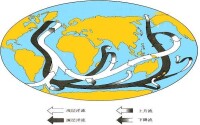
(圖)大洋環流
大洋環流的演變大洋環流的演變是控制古氣候變化(特別是新生代氣候變冷)的基本因素之一,而環流的變遷又受到板塊運動和陸塊間各海水通道開閉的支配。
二疊紀、三疊紀期間(距今約2億多年前),地球上僅有一個聯合古陸,周圍是統一的泛大洋(古太平洋),南、北半球大洋中分別存在單一的巨大環流。泛大洋西緣較暖,東緣較冷,南北向的溫度梯度反而不及東西向的梯度大。晚三疊世,聯合古陸開始解體。侏羅紀北美開始與南美、非洲分裂,形成北大西洋,在閉塞環境中接受了蒸發岩沉積。晚侏羅世至白堊紀,北大西洋東通特提斯海,西南經中美海道連接太平洋,形成了自東向西環繞全球的赤道環流。當時太平洋在南、北半球均發育亞熱帶反氣旋環流和副極地氣旋環流。早白堊世南大西洋和印度洋張開,但初期南、北大西洋並不連通,在南大西洋形成大量蒸發岩。1.1~0.9億年前,南、北大西洋之間出現表層水交流;在0.7億年前,進一步出現深層水交流。當時北大西洋與北冰洋仍然彼此隔絕,直至新生代初期,北大西洋才開始與北冰洋連通。隨著北美大陸西漂與亞洲靠攏,自晚白堊世開始,北冰洋與太平洋之間的深水交流終止。
新生代以來大洋環流的變遷,主要表現在以下三方面:
① 赤道環流的減弱和中斷。(i)4000 萬年前印度與亞洲主體匯合,使赤道環流局限於從阿拉伯以北的狹窄水道通過;1800萬年前,非洲-阿拉伯與歐亞板塊的碰撞和中東造山運動使特提斯海道最終封閉,大西洋-地中海與印度洋之間的交流終止。(ii)新生代晚期,隨著澳大利亞北移,印度尼西亞海道關閉,赤道印度洋與赤道太平洋之間的深水交流受阻。(iii)上新世晚期(約350萬年前)巴拿馬地峽形成,切斷了赤道太平洋與赤道大西洋之間的水體交流。
② 環南極洋流的形成。(i)漸新世最早期,隨著澳大利亞與南極洲進一步分離(最初分裂在始新世初),塔斯馬尼亞以南的南塔斯曼隆起與南極大陸分開,沿新裂開的塔斯馬尼亞海道,印度洋海水流入太平洋。(ii)漸新世晚期或中新世初期,南美洲與南極洲之間的德雷克深水通道張開,完整的環南極洋流最終形成。
③ 南極底層水的形成。漸新世最早期,隨著南極地區冰川和海冰的發展,寒冷的高鹽度表層海水下潛,形成南極底層水(冷水圈環流),從而導致大洋底層水富氧,海底侵蝕作用增強,沉積間斷及再沉積作用盛行。至晚第三紀,統一的赤道環流已不復存在,環南極洋流十分強勁,開始出現類似於現代的大洋環流特點。
新生代氣候變冷過程 對深海岩心的氧同位素(18O/16O)分析表明,白堊紀以來氣候有變冷趨勢(圖2)。新生代變冷過程的特點是:高緯地區明顯變冷,低緯地區不甚顯著;海洋深層水明顯變冷,表層水不甚顯著;變冷並不是平穩的漸變過程,在溫度下降的總趨勢上疊加著幾次急劇的氣候波動。
始新世末、中中新世和晚上新世為三次急劇變冷期:
① 始新世末期事件。距今3700萬年前,海洋底層水溫驟降4~5°C,南極周圍首次形成大規模海冰,開始出現寒冷的南極底層水。南極大陸上已有冰川,但可能尚未成為巨大冰蓋。有孔蟲等生物蒙受沉重打擊,漸新世初期海洋生物的分異度極低。這些情況的發生可能與塔斯馬尼亞海道的張開有關。南印度洋高緯海域的表層冷水,經塔斯馬尼亞海道注入南極羅斯海域,取代了南下的東澳大利亞暖流,從而觸發了南極地區的冰凍。
② 中中新世南極冰蓋形成。大約1400萬年前,南極大陸形成冰蓋,南大洋海冰進一步擴展(當時北半球仍無冰川),冰載沉積物分佈甚廣,硅質生物沉積帶向北推展。這一時期的海洋進一步變冷與環南極洋流的形成和強化有關,它使南極水體無法與低緯度溫暖水體交流,南極大陸在一定程度上處於熱隔絕狀態。中新世中期冰鳥-法羅海脊沉沒,北冰洋-挪威海水注入北大西洋,北大西洋深層水長驅南下,然後上升,加入環南極洋流,這可能為南極大陸帶來了形成冰蓋所必需的降水。
③ 晚上新世北半球冰蓋形成。大約300~240萬年前,北大西洋和北太平洋出現冰載沉積物,標誌北半球冰蓋的形成,由此開始出現冰期-間冰期氣候旋迴。北半球生成冰蓋,可能與巴拿馬地峽形成,墨西哥灣流強化有關。灣流為形成冰蓋帶來了水汽。
第四紀海洋 據氧同位素的資料研究,第四紀冰期與間冰期頻繁更迭,最突出的變動周期約10萬年,其上還疊加著約4萬年和2萬年的周期。這些周期長短分別與地球軌道偏心率、自轉軸傾角及歲差的變動周期相當,因此,冰期-間冰期旋迴可由地球軌道變動說(米蘭科維奇理論)作出比較合理的解釋。隨著冰期、間冰期的交替,反覆發生海退與海侵,海面變動幅度可達100米左右;深海沉積物的碳酸鹽丰度發生周期性變化,構成碳酸鹽溶解旋迴;氣候帶(特別是極峰線)南、北遷移,冰載沉積物分佈區時進時退。至第四紀後半期,氣候變動的幅度達到最大。距今70萬年以來北極冰蓋增厚。隨著溫度梯度和大洋環流的增強,南大洋及其他一些海域生物生產力升高,至第四紀晚期達到新生代最高值。在冰期,冰蝕作用強盛,海面下降導致河流侵蝕作用增強,氣候乾冷導致沙漠擴展,卷揚起大量泥砂,從而使大洋中陸源沉積速率明顯增大。
據“遠期氣候調查、測繪和預報”(CLIMAP)的結果表明:18000年前為末次冰期鼎盛期,當時冰蓋廣達陸地面積的三分之一,厚可達3公里。海平面至少比現代低85米。18000年前的大洋表層水溫分布圖顯示:極峰線向赤道偏移,海水溫度低,表層海水溫度比現在低2~3°C;溫度梯度高,尤以北大西洋和南大洋最為顯著;沿非洲、澳大利亞和南美洲西岸的東部邊界流強盛,伴隨著冷水向赤道方向擴展;赤道上升流和沿岸上升流也很活躍。近10000多年來,溫度升高,冰川退縮,海面回升。目前地球正處在間冰期。
深海鑽探和同位素地球化學等方面的古海洋學研究,揭示了大洋演化中的一系列重大事件。除上述大洋環流與氣候變冷過程中的事件外,重要者有白堊紀中期缺氧事件,白堊紀末生物絕滅事件和中新世末地中海變幹事件。
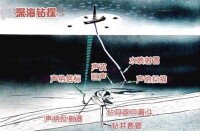
(圖)深海鑽探
白堊紀末生物絕滅事件 距今6600萬年以前,恐龍、菊石、浮遊有孔蟲、超微浮游生物和箭石等大批絕滅。據統計約有二分之一的屬消亡了。許靖華等通過深海鑽探岩心的研究,發現白堊-第三系界面處,CaCO3含量極低,顯示鈣質浮游生物數量驟減,碳酸鹽補償深度一度急劇上升;18O/16O比值陡降表明溫度突然上升,13C/12C比值聚降,也反映了大洋化學和溫度的急劇變化。關於這一事件的起因,眾說紛紜。諸如氣候變化、海面變動、地磁場倒轉、火山爆發等說法,難以解釋生物絕滅的突發和全球性質。鑒於界面粘土層中銥含量極高,而隕石中富含銥,一些學者主張星體隕落說。大隕星撞擊地球釋放出的巨大能量,導致溫度突然升高,臭氧層遭到破壞;衝擊作用掀起的大量塵埃遮蔽陽光,植物的光合作用受阻,可導致食物鏈中斷而危及一系列生物。
中新世末地中海變幹事件 深海鑽探揭示,地中海海底埋藏著中新世末期(墨西拿期,距今約 500多萬年前)的蒸發岩。蒸發岩厚可達2~3公里,總體積100萬立方公里,致使大洋水的鹽度降低2‰,這一事件使地中海的海洋生物陷於絕境,故稱地中海鹽度危機,也叫墨西拿事件。1800萬年前中東造山運動切斷了地中海與印度洋之間的聯繫,地中海成為西通大西洋的內陸海。中新世末期,由於板塊匯聚,地中海西端海峽變淺,加之海面下降,使地中海成為隔絕狀態。蒸發岩的上覆和下伏地層均屬半遠洋沉積,底棲有孔蟲的古深度分析表明,地中海在變干前後有類似於現代的深海環境。硬石膏、疊層石和乾裂等現象說明這一深海盆地曾經乾涸。乾涸時期,周圍大陸的河谷深切至現代海面以下數百米,河流在裸露的大陸坡上刻蝕出深邃的峽谷。至上新世初期,直布羅陀海峽打開,大西洋水再度進入地中海。
[1]大科普網 http://www.ikepu.com/chemistry/chemistry-index.htm