共找到2條詞條名為杜鵑蘭的結果 展開
- 蘭科杜鵑蘭屬植物
- 中藥
杜鵑蘭
蘭科杜鵑蘭屬植物
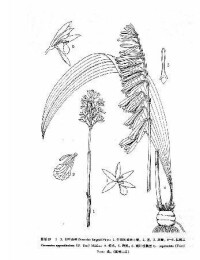
杜鵑蘭(學名:Cremastra appendiculata (D. Don) Makino):假鱗莖卵球形或近球形,密接,有關節,外被撕裂成纖維狀的殘存鞘。葉通常1枚,生於假鱗莖頂端,狹橢圓形、近橢圓形或倒披針狀狹橢圓形。花葶從假鱗莖上部節上發出,近直立;總狀花序長10-25厘米,具5-22朵花;花常偏花序一側,多少下垂,不完全開放,有香氣,狹鐘形,淡紫褐色;花瓣倒披針形或狹披針形,向基部收狹成狹線形,長1.8-2.6厘米,上部寬3-3.5毫米,先端漸尖;唇瓣與花瓣近等長,線形,上部1/4處3裂。蒴果近橢圓形,下垂。花期5-6月,果期9-12月。
生於林下濕地或溝邊濕地上,海拔500-2900米。分佈於中國、尼泊爾、不丹、錫金、印度、越南、泰國和日本。此花有栽培,具有較高的園藝價值。

杜鵑蘭
杜鵑蘭具有卵球形的假鱗莖,一般有鵪鶉蛋大小,外面布滿了纖維狀的殘存鞘。僅有的一枚橢圓形葉片就從假鱗莖的頂端生出,約有成人手掌一般大。葉片無力的趴在地面上,上面布滿了黃色的斑點,彷彿得了重病一般。或許,只有看那伸出的一根花葶才知道,她還有生命力,還在履行著作為生物體的本能——繁殖。總狀花序有20厘米長,10餘朵黃色的花兒懸掛在花葶上。花萼細長,先端尖;花瓣倒披針形,唇瓣與其近等長,線形。每年的5 ~ 6月份開花,9 ~ 12月份結出近橢圓形的蒴果。
杜鵑蘭不僅有著暗藏的驚艷相貌,還有著豐富的“內涵”。假鱗莖可入葯,性味辛、澀,可清熱解毒,潤肺止咳,活血止痛,對癰腫、跌打損傷和蚊蟲叮咬等具有良好的治療效果。
國內分佈:產山西南部(介休、夏縣)、陝西南部、甘肅南部、江蘇、安徽、浙江、江西(廬 山)、台灣、河南、湖北、湖南、廣東北部(乳源)、四川、貴州、雲南西南部至東南部 (鳳慶、西疇)和西藏。生於林下濕地或溝邊濕地上,海拔500--2900米。
國外分佈:尼泊爾、不 丹、錫金、印度、越南、泰國和日本也有分佈。模式標本采自印度。
杜鵑蘭
多少 =下垂,不完全開放,有香氣,狹鐘形,淡紫褐色;萼片倒披針形,從中部向基部驟然收狹而成近狹線形,全長2––3厘米,上部寬3.5––5毫米,先端急尖或漸尖;側萼片略斜 歪;花瓣倒披針形或狹披針形,向基部收狹成狹線形,長1.8––2.6厘米,上部寬3— 3.5毫米,先端漸尖;唇瓣與花瓣近等長,線形,上部1/4處3裂;側裂片近線形,長 4––5毫米,寬約1毫米;中裂片卵形至狹長圓形,長6––8毫米,寬3—5毫米,基部 在兩枚側裂片之間具1枚肉質突起;肉質突起大小變化甚大,上面有時有疣狀小突起;蕊柱細長,長1.8––2.5厘米,頂端略擴大,腹面有時有很狹的翅。蒴果近橢圓形,下垂,長2.5––3厘米,寬1—1.3厘米。花期5––45月,果期9—12月。
本種提示此種是一個廣布種,唇瓣側裂片的形狀和中裂片基部肉質突起均有較大變化,蕊柱上部的狹翅也有類似情況。一些學者將唇瓣上肉質突起較小而平滑,以及蕊柱上部腹面 有狹翅的類型分出,另立為種或變種。但根據中國標本,此種特徵不僅有過渡,而且有交叉,難以分割,故我們不予劃分,仍視為同一個種。
Lindl,Gen.Sp.Orch,Pl.172.1833;I.D.Lund in Nord.J. Bot.8(2):197.1987.地生草本,地下具根狀莖與假鱗莖。假鱗莖球莖狀或近塊莖狀,基部密生多數纖維 根。葉1––2枚,生於假鱗莖頂端,通常狹橢圓形,有時有紫色粗斑點,基部收狹成較 長的葉柄。花葶從假鱗莖上部一側節上發出,直立或稍外彎,較長,中下部具2—3枚 筒狀鞘;總狀花序具多朵花;花苞片較小,宿存;花中等大;萼片與花瓣離生,近相 似,展開或多少靠合;唇瓣下部或上部3裂,基部有爪並具淺囊;側裂片常較狹而呈線 形或狹長圓形;中裂片基部有l枚肉質突起;蕊柱較長,上端略擴大,無蕊柱足;花粉團4個,成2對,兩側稍壓扁,蠟質,共同附著於粘盤上。本屬模式種:Cremastra appendiculata(D.Don) Makino (Cymbidium appendicula– tum D.Don) · 全屬僅2種,分佈於印度北部、尼泊爾、錫金、不丹、泰國、越南、日本和中國秦 嶺以南地區。中國2種均產。
Camus,Bergon et A.A.Camus E.G.Carnus,Bergon et A.A.Calnus,Monogr,Orch,Eur, Afr.Septentr.Etc.376.1908.地生植物,地下具球莖狀或塊狀假鱗莖;假鱗莖非綠色,具節,以匍匐根狀莖相連接,頂端具1——2枚葉。葉草質至紙質,具或不具關節,通常有葉柄。花葶側生或頂生,不分枝;總狀花序具數花或多花,有時減退為單花;花小至中等大,扭轉;萼片與花瓣離生;唇瓣常多少3裂;有距或無距;蕊柱長或中等長,通常無蕊柱足;花粉團4個,成2對,蠟質,具或不具粘盤柄,通常有粘盤;蕊喙小;柱頭凹陷。本亞族模式屬:Calypso Salisb.共含8屬,分佈於北溫帶和東亞。中國有5屬。
地生、附生或腐生草本,具根狀莖、塊莖或假鱗莖。葉莖生或基生,摺扇狀、對摺或具網狀脈,草質至革質。花序(或花葶)頂生或側生;花通常兩側對稱,有特化的唇瓣,少有例外,一般扭轉,具1室子房;蕊柱通常具1枚能育雄蕊;花粉粘合成團塊,稱花粉團;花粉團2––8個,粒粉質或蠟質,其一端常伸出花粉團柄並連接於粘盤,也有直接連接於粘盤或通過粘盤柄連接於粘盤的,較少既無花粉團柄又無粘盤柄與粘盤的;柱頭1或2,凹陷或凸出;蕊喙通常存在,大或小。果實通常為蒴果;較少呈莢果狀。種子通常兩端有延長的翅,較少無翅並有厚的外種皮。本亞科模式屬:OrchisL.共約690餘屬,全球均有分佈。中國有4個族,42個亞族共167屬。

杜鵑蘭
此時2枚側萼片基部常著生於蕊柱足上,形成囊狀結構,稱萼囊;花粉通常粘合成團塊,稱花粉團,花粉團的一端常變成柄狀物,稱花粉團柄;花粉團柄連接於由蕊喙的一部分變成固態粘塊即粘盤上,有時粘盤還有柄狀附屬物,稱粘盤柄;花粉團、花粉團柄、粘盤柄和粘盤連接在一起,稱花粉塊,但有的花粉塊不具花粉團柄或粘盤柄,有的不具粘盤而只有粘質團。果實通常為蒴果,較少呈莢果狀,具極多種子。種子細小,無胚乳,種皮常在兩端延長成翅狀。科的模式屬:OrchisL.全科約有700屬20000種,產全球熱帶地區和亞熱帶地區,少數種類也見於溫帶地區。中國有171屬1247種以及許多亞種、變種和變型。蘭科是一個進化而複雜的科,至今對科內各類群間親緣關係的了解仍然是十分初步的。在目前已發表的許多蘭科系統中,以R.Dressier(1981,1993)系統較為全面,但該系統對鳥巢蘭族(ridb.Neottieae)的處理是有欠缺的。幸而C.Seidenfaden等人在“TheOrehidsofIndochina”(OperaBot.114:5––502.1992)和“TheOrehidsofPeninsularMalaysiaandSingapore”(1992)中有較合理的處理。
基本上採用了後者的框架,但將等級作了調整,而對亞族的組成則更多地參考Dressler系統(PhylogenyandClassifica–tionoftheOn:hidFamily,1993)。本書將蘭科分為3個亞科,蘭亞科之下分4個族。其中蘭族(Orchideae)與萬代蘭族(Vandeae)是比較自然的類群,而鳥巢蘭族(Neottieae)與樹蘭族(Epidendreae)則尚存在不少問題。在許多近代蘭科系統中,上述的4個族均被作為亞科,在亞科之下又有大量的族,在族之下則是亞族。應當說,大多數亞族的建立是比較客觀的。但族的組合則含有相當多的主觀性,甚至是勉強的拼湊。因此,避繁就簡,在蘭亞科之下只承認4個族,族之下則包含較多的亞族。這樣不僅簡化了層次,而且更為客觀和實用。當然,這也只是權宜之計,有些問題仍然未能解決,如鳥巢蘭族與樹蘭族的特徵交叉過多,有些亞族較為龐雜等。這裡就不作詳細討論了。這裡要說明的是,所提供的“亞科與族的檢索表”和“蘭亞科分亞族檢索表”均系根據國產種類編寫而成的,未必完全符合族與亞族的全面特徵,在使用時務請加以注意。
(1)採用杜鵑蘭蒴果種子和假球莖上的芽眼作為外植體,其中蒴果種子進行無菌播種培養,萌發形成原球莖;假球莖芽眼經消毒後進行接種培養,獲得原球莖;
(2)通過原球莖在培養基里的增殖,將其增殖3~7倍;
(3)將增殖所得的原球莖分化成苗並促其生根,獲得完整的再生植株;
(4)將再生植株栽種在蛭石苗床或營養缽生長、過渡。或將增殖的根狀莖直接栽種在蛭石里生根發芽得到種苗。將種苗栽種到準備就序的土壤中,並進行栽培管理,得到人工栽培的杜鵑蘭假球莖。採用本方法對杜鵑蘭進行種苗繁殖,有效地解決了杜鵑蘭繁殖困難和種苗缺乏的問題,可實現杜鵑蘭的規模化、工廠化生產。

杜鵑蘭
2、揭示了菌根真菌動態變化特徵及其與植物生長發育的關係利用新建立的單菌絲團分離方法,對三種蘭科植物不同生長期、根中菌根真菌的縱向和橫向變化的研究,發現植物根形成早期的優勢菌生長快、氣生菌絲髮達,後期優勢菌生長慢、氣生菌絲不發達。菌根真菌由根尖向基部及由根表皮向內,生長快、氣生菌絲髮達的菌根真菌比例下降,生長較慢、氣生菌絲不發達的菌根真菌比例增加。
3、揭示了菌根真菌生理生化特徵及其與植物生長發育的關係通過對三種蘭科植物菌根真菌對不同碳源、氮源的利用能力,產B族維生素和赤霉素的能力、是否具有纖維素酶和多酚氧化酶酶活的研究,發現營養生長階段的優勢菌根真菌纖維素酶活較弱,多酚氧化酶活陽性或弱陽性,形成的B族維生素的量較高,澱粉、蔗糖、麥芽糖和無機氮的利用能力較強。生殖生長期優勢菌纖維素酶活較強,多酚氧化酶活弱或無,形成的B族維生素的量低,對纖維素、澱粉和有機氮的利用能力強。
4、從菌根營養的角度揭示了菌根真菌提高組培苗移栽成活率的內在原因,提出了“三個營養庫”的假說和杜鵑蘭組培苗移栽成功的“三個策略”通過對三種蘭科植物菌根真菌與杜鵑蘭各生長期的組培苗共培養,發現杜鵑蘭各生長階段的促生菌是此階段的優勢菌,同時發現不同生長期的組培苗移栽成活率不同,已分化根狀莖、帶根狀莖的幼苗、帶球莖的幼苗成活率高,而無根狀莖或球莖的幼苗移栽成活率低,同時發現菌根真菌在組培苗移栽過程中主要起保護根及提高根的活力的作用。據於此提出了杜鵑蘭組培苗移栽成活密切相關的“三個營養庫”的假說,即根狀莖、葉片和球莖,組培苗移栽成功就在於成功實現“根狀莖營養庫”到“葉片光合營養庫”,再到“球莖營養庫”的轉變,並以此為指導,總結出杜鵑蘭組培苗移栽成功的“三個策略”,即選擇根狀莖、帶根狀莖的分化苗或帶球莖的分化苗,進行菌根化育苗,利用菌根真菌的保根作用、根狀莖和球莖的營養庫作用,使組培苗成功實現向“光合營養庫”的轉變,並最終建立可順利渡過休眠期的“球莖營養庫”。
5、建立了菌根真菌應用於組培苗移栽的模型通過總結蘭科植物菌根真菌動態變化特點、菌根真菌生理生化特性、蘭科植物本身特性等的研究結果,形成了蘭科植物菌根真菌應用於組培苗移栽的模型,選擇帶有營養貯存器官的蘭科植物組培苗與根形成早期的優勢菌根真菌,進行菌根化培養,促進生根及提供菌根營養,待共生關係形成並建立了穩定的光合作用后,施用營養生長期優勢菌,促進蘭科植物光合產物轉化貯存和保持根的高活力,實現蘭科植物組培苗的成功栽培。
6、完成三種蘭科植物菌根真菌的分類鑒定採用形態鑒定、酶活特性測定和ITS序列系統發育分析方法,完成三種蘭科植物菌根真菌的分類鑒定,較為系統地提出了蘭科植物共生絲核菌的分類鑒定方法及相關屬種的分類檢索表,並發現Epulorhiza屬中的第三個種群,即未定名的新屬Newgenus,該屬的最大特點是生長很慢、菌落邊緣不整齊、不形成或形成形狀不規則的念珠狀細胞,澱粉利用能力強,是蘭科植物光合產物貯存、轉化及利用相關的重要菌根真菌之一。研究中提出的“三個營養庫”的假說,將豐富蘭科植物菌植共生理論,提出的蘭科植物共生絲核菌的分類方法、相關屬種檢索表、提出的新屬,將豐富蘭科植物共生絲核菌的分類鑒定體系。本研究建立的單菌絲團分離方法、提出的杜鵑蘭組培苗移栽的“三個策略”及建立菌根真菌應用於組培苗移栽的模型,為實現杜鵑蘭、獨蒜蘭和雲南獨蒜蘭組培苗的成功移栽,實現其產業化提供理論和技術支撐。
