基因連鎖
基因連鎖
基因的連鎖和交換定律指是在進行減數分裂形成配子時,位於同一條染色體上的不同基因,常常連在一起進入配子;在減數分裂形成四分體時,位於同源染色體上的等位基因有時會隨著非姐妹染色單體的交換而發生交換,因而產生了基因的重組。應當說明的是,基因的連鎖和交換定律與基因的自由組合定律並不矛盾,它們是在不同情況下發生的遺傳規律:位於非同源染色體上的兩對(或多對)基因,是按照自由組合定律向後代傳遞的,而位於同源染色體上的兩對(或多對)基因,則是按照連鎖和交換定律向後代傳遞的。
在1906年,科學家貝特生等在研究香豌豆的兩對相對性狀時,發現同一親本的兩種性狀,在雜交後代中,比較多地連在一起出現,並不按照孟德爾自由組合規律的比例發生分離,這使他們感到非常困惑,甚至對孟德爾的遺傳規律產生懷疑。美國的遺傳學家摩爾根和他的同事用果蠅做實驗材料,進行了大量的遺傳學研究,終於解開了人們心中的疑團,這不僅證實了孟德爾的遺傳規律的正確性,並且豐富發展了關於兩對(或兩對以上)基因的遺傳理論,提出了遺傳的第三個規律----基因的連鎖互換規律。
兩對(或兩對以上)的等位基因位於同一對同源染色體上,在遺傳時位於同一個染色體上的不同(非等位)基因常常連在一起不相分離,進入同一配子中。
具有連鎖關係的兩個基因,其連鎖關係是可以改變的。在減數分裂時,同源染色體間的非姐妹單體之間可能發生交換,就會使位於交換區段的等位基因發生互換,這種因連鎖基因互換而產生的基因重組,是形成生物新類型的原因之一。
摩爾根等人用純種灰身長翅果蠅與純種黑身殘翅果蠅交配,他們看到子一代(F1)都是灰身長翅的,由此可以推出,果蠅的灰身(B)對黑身(b)是顯性;長翅(V)對殘翅(v)是顯性。所以,純種灰身長翅果蠅的基因型與純種黑身殘翅果蠅的基因型應該分別是(BBVV)和(bbvv).F1的基因型應該是(BbVv)(如圖).
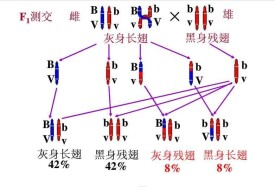
摩爾根又讓F1的雄果蠅(BbVv)與雙隱性類型的雌果蠅(bbvv)測交,按照自由組合定律,測交後代中應該出現4種不同的類型,即灰身長翅,灰身殘翅,黑身長翅,黑身殘翅,並且它們之間的數量比應該為1:1:1:1.但是,測交的結果與原來預測的完全不同,只出現兩種和親本完全相同的類型:灰身長翅(BbVv)和黑身殘翅(bbvv),並且兩者的數量各佔50%.很明顯,這個測交的結果是無法用基因的自由組合定律來解釋的. 讓F1的雌果蠅與黑身殘翅雄蠅交配,測交後代表現型及其數目是:21灰身長翅(42%),4灰身殘翅(8%),21黑身長翅(42%),8黑身殘翅(8%)。
為什麼會出現上述試驗結果呢 摩爾根認為果蠅的灰身基因和長翅基因位於同條染色體上,可以用來表示(如圖);黑身基因和殘翅基因也位於同一條染色體上,可以用來表示。所以,當兩種純種的親代果蠅交配后,F1的基因型BbVv,應該表示為,表現型是灰身長翅。這樣,在F1雄果蠅產生配子時,原來位於同一條染色體上的兩個基因(B和V,b和v)就不能分離,而是連在一起向後代傳遞。因此,當F1雄果蠅與黑身殘翅的雌果蠅交配后,只能產生灰身長翅()和黑身殘翅()兩種類型,並且這兩者的數量各占 50%.像這樣,位於一對同源染色體上的兩對(或兩對以上)等位基因,在向下一代傳遞時,同一條染色體上的不同基因連在一起不相分離的現象,叫做連鎖。在上述雄果蠅的測交試驗中,由於只有基因的連鎖,沒有基因之間的交換,因此,這種連鎖是完全連鎖.在完全連鎖遺傳中,後代只表現出親本類型.
摩爾根等人還做了另一組試驗,他們讓子一代(F1)的雌果蠅(BbVv)與雙隱性類型的雄果蠅(bbvv)測交,所得的結果如圖所示。從圖中所示的結果可以看出,F1與雙隱性類型測交,雖然測交後代的表現型與基因自由組合定律中測交的結果一樣,也是4種類型;灰身長翅,灰身殘翅,黑身長翅和黑身殘翅,但是,它們之間的數量比並不符合基因的自由組合定律中的1:1:1:1,而是與親本表現型相同類型的比例很大(佔總數的84%);與親本表現型不同類型的比例很小(佔總數的16%).
為什麼會出現上述的試驗結果呢 摩爾根認為,位於同一條染色體上的兩個基因的連鎖關係有時是可以改變的(如圖).在細胞進行減數分裂形成配子的過程中(即出現四分體時),如果同源染色體中,來自父方的染色單體與來自母方的染色單體相互交換了對應部分,在交換區段上的等位基因就會發生交換,這種交換可以產生新的基因組合。所以測交后,在子代產生了與親代表現型相同類型的同時,也產生了與親代表現型不同的新類型。但是,為什麼測交後代的數量比不是1:1:1:1呢 這是因為F1在形成配子時,大部分配子中的同一條染色體上的這兩個基因是連鎖的,因而生成的配子和配子特別多(各佔42%),只有一小部分配子中的兩個基因因為交換(交叉點正好位於基因B與V, b與v的中間)而產生了新的組合,因而生成的配子和配子很少(各佔8%).因此,F1與雙隱性類型測交,就產生了這樣的結果:灰身長翅佔42%,黑身殘翅佔42%,灰身殘翅佔8%,黑身長翅佔8%.在上述雌果蠅的測交試驗中,由於基因在向下一代傳遞的過程中,不僅有連鎖,還出現了交換,因此,這種遺傳是不完全連鎖遺傳.
基因的連鎖和交換定律的實質:
基因連鎖和互換規律的實質:
位於同一染色體上的不同基因,在減數分裂過程形成配子時,常常連在一起進入配子;在減數分裂的四分體時期,由於同源染色體上的等位基因隨著非姐妹染色單體的交換而發生互換,因而產生基因的重組。
三個規律的聯繫:
基因的自由組合規律和基因的連鎖互換規律是建立在基因的分離規律的基礎上的,生物形成配子時,在減數第一次分裂的過程中,同源染色體上的等位基因都要彼此分離。在分離之前,可能發生部分染色體的交叉互換。在同源染色體分離的基礎上,非同源染色體上的非等位基因又進行自由組合,從而形成各種組合的配子。可見三大規律在配子形成過程中相互聯繫、同時進行、同時作用。
基因的連鎖和交換定律,在動植物育種工作和醫學實踐中都具有重要的應用價值。如果不利的性狀和有利的性狀連鎖在一起,那就要採取措施,打破基因連鎖,進行基因互換,讓人們所要求的基因連鎖在一起,培育出優良品種來。
在育種工作中,人們根據育種目標選配雜交親本時,必須考慮基因之間的連鎖關係。如果幾個有利性狀的基因連鎖在一起,這對育種工作就很有利。例如,大麥抗稈鏽病與抗散黑穗病的基因就是緊密連鎖的,在育種中只要注意選擇大麥抗稈鏽病的植株,也就等於同時選擇了抗散黑穗病的植株,達到一舉兩得,提高選擇效率的目的。但是如果不利性狀與有利性狀的基因連鎖在一起,就要採取措施打破基因連鎖,促成基因交換,讓人們所需要的基因重組在一起,從而培育出優良品種來。例如,有兩個大麥品種:一個是矮稈抗倒伏但不抗鏽病的品種,另一個是高稈易倒伏但抗鏽病的品種。每一個品種中控制這兩個性狀的基因都位於同一條染色體上。經過雜交,F2會出現四種類型的後代,其中由於基因交換而出現的矮稈抗倒伏同時又抗鏽病的類型就是符合需要的類型,經過進一步培育和大量繁殖就可以成為良種,其他不符合需要的類型應該淘汰。由此可見,通過基因交換產生的新類型能夠為育種工作提供原始材料.
在醫學實踐中,人們可以利用基因的連鎖和交換定律,來推測某種遺傳病在胎兒中發生的可能性。例如,有一種叫做指甲髕骨綜合症的人類遺傳病.患者的主要癥狀是指甲發育不良,髕骨缺少或發育不良。這種病是一種顯性遺傳病,致病基因(用兩個大寫字母NP表示)與ABO血型的基因(IA,IB或i)位於同一條染色體上。在患這類疾病的家庭中,NP基因與IA基因往往連鎖,而NP的正常等位基因np與IB基因或i基因連鎖,又已知NP和IA之間的重組率為10%.由此可以推測出,患者的後代只要是A型或AB型血型(含IA基因),一般將患指甲髕骨綜合症,不患這種病的可能性只有10%.因此,這種病的患者在妊娠時,應及時檢驗胎兒的血型,如果發現胎兒的血型是A型或AB型,最好採用流產措施,以避免生出指甲髕骨綜合症患兒