AMPA受體
AMPA受體
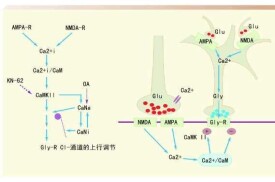
AMPA 受體(α-氨基-3-羥基-5-甲基-4-異惡唑丙酸受體,AMPAR)介導中樞神經系統快速興奮性突觸傳遞,其在突觸后膜的動態表達與長時程增強、長時程抑制的誘發和維持有關,參與調節學習記憶活動。AMPAR在β-澱粉樣蛋白作用下的過度胞吞和裂解致其在突觸后膜缺失,可致突觸損傷和功能障礙,與阿爾茨海默病早期認知障礙密切相關。AMPAR還參與谷氨酸介導的興奮性損傷,Ca2+通透性AMPAR亞型的過度激活能導致阿爾茨海默病神經元的功能障礙甚至死亡。此外,AMPAR還參與tau蛋白的異常磷酸化,與神經原纖維纏結的形成有關。因而突觸后膜AMPA受體數目和功能異常可能是導致阿爾茲海默病發生的重要環節。
AMPA受體是由GluR1-4 (GluRA-D) 4個亞基組成的四異聚體,其形成起始於粗面內質網各個亞基的合成 ,每個亞基都有1個大的N端、3個跨膜區域、1個形成孔的髮夾結構和1個位於胞質側的C端。成年海馬AMPA受體主要由GluR和GluR2或 G luR2和 G luR3 所組成的異聚體構成 ,而GluR和GluR2組成的受體只存在於幼年海馬和其他成熟腦區[2 ]儘管各個亞基的胞外結構和跨膜區非常相似 ,但他們的胞質側 C端卻不盡相同, GluR1, GluR4 和少見的GluR2L 相似 ,有較長的 C端 ,而多見的 GluR2, GluR3和少見的GluR4c具有較短的 C端 ,各個亞基通過 C端和不同的胞內蛋白相互作用突觸后緻密區(postsynaptic density zone ,PDZ)是突觸后膜結構的骨架成分 , 大量不同類型的細胞實驗已證明突觸后緻密區在將膜蛋白定向或聚集到特定亞細胞區域的過程中發揮著重要作用。多數 AMPA 受體反應蛋白有相應的單一或多元 PDZ,目前已知 ,G luR1對應經典的ⅠPDZ配體, 而GluR2G luR3和 G luR4c 對應經典的ⅡPDZ 配基。由於 G luR4 和G luR2L 的 C端有多種形式 ,所以它們是否與經典的 PDZ區域蛋白反應尚不清楚。四聚體的 AMPA受體由相應的 4 個亞基交互排列圍繞形成受體通道[3 ],其主要功能是介導 Na+和 K+及少量 Ca2 +內流。其中 ,限制通道鈣離子電導的主要因素— — GluR2在受體運動早期位於粗面內質網內部 ,而控制GluR2移出粗面內質網的結構成分位於胞質側的 C端和髮夾結構形成的孔頂端。另外 ,在到達突觸之前 ,四聚體要在高爾基體進行糖基化反應。
近年來,靜寂突觸的存在和LTP過程中 AMPA受體快速的突觸插入已被大量實驗證實,而產生這一現象的重要前提是在突觸附近存在可被利用的非突觸 AMPA 受體池。用顯微技術觀察谷氨酸受體在突觸及突觸附近的分佈發現,除突觸膜上存在有突觸性的AMPA受體外,在樹突的表面和胞質中也存在大量的非突觸性 AMPA 受體。
而且 ,非突觸性AMPA受體數目要遠遠多於突觸性AMPA受體。用膠體金技術研究表明,在海馬 CA1 區錐體細胞突觸膜上,NMDA受體遠比AMPA受體的分佈廣泛。