偶蹄目
哺乳動物中的一個目
偶蹄目(學名:Artiodactyla )也被表述為 鯨偶蹄目(Cetartiodactyla ),是哺乳動物的一個大型分支。陸生種類四足通常各有四趾或二趾,趾甲特化為鞘狀的角質蹄,因每足的蹄甲數為偶數(四或二),故稱偶蹄目。
偶蹄目源自5000萬年前始新世的踝節目,演化至今已成為哺乳類中最繁盛的家族之一。現存300餘種,可分4個亞目:胼足亞目(駱駝和羊駝類)、豬形亞目(豬和西猯)、反芻亞目(牛、羊、鹿 等)、鯨河馬亞目(鯨豚類與河馬) 。
近年來古生物學和遺傳學研究都強烈支持鯨類起源於偶蹄目,由陸地上的原始偶蹄類進入水中生活演化而來。DNA序列檢測顯示,與河馬親緣關係最近的動物是鯨類,而非其他陸生偶蹄類,同時河馬、鯨類比豬和駱駝更接近反芻亞目,鯨類顯然是偶蹄目不可分割的一部分。傳統的形態學分類將鯨豚類單列為鯨目(Cetacea),獨立於偶蹄目之外,這使得偶蹄目被定義為無效的並系群,因此現行分類將鯨目歸入偶蹄目,旨在構建一個完整的、有效的單系群。
陸生偶蹄類為植食性或雜食性,以植物莖葉、根、果實和小型動物、殘骸腐肉等為食;水生偶蹄類(鯨)為肉食性,捕食魚蝦、烏賊甚至海獸。
偶蹄目由19世紀英國生物學家理查德·歐文(RichardOwen)在1848年命名,“Artiodactyla”來自古希臘文 ἄρτιος(ártios,意為“偶數”)和δάκτυλος(dáktylos,意為“手指,腳趾”)。
偶蹄動物由古新世的踝節目動物進化而來,約從始新世開始分化,中新世和上新世是其進化的重要時期。早期偶蹄目中雖然有些較成功的類群,但總體上並不繁盛,中新世以來生態位日趨顯著,到現代則佔據壓倒性的絕對優勢,無論種類、數量還是分佈上,均遠遠超過其他現存的有蹄類。
早期偶蹄動物形似現代的鼷鹿:小巧、腿短、取食植物莖葉。4600萬年前的始新世晚期,偶蹄目的4個亞目已經開始分化,但它們並未佔據生態主導地位,時值奇蹄目的興盛期,偶蹄目只能佔據一些邊緣生態位艱難繁衍。然而,偶蹄目在此時開始進化出複雜的消化系統,從而能夠依靠劣質食物為生。
始新世開始出現草。中新世全球氣候變得乾燥少雨,大量雨林枯亡,草原開始發育,並向全球蔓延開來。草本身是一種很難消化的食物,而擁有複雜消化系統的偶蹄目能有效利用這種粗糙、低營養的食物。新近紀以後偶蹄動物取代了奇蹄動物的生態位,成為食草動物的主體。
分子系統學研究表明,鯨目起源於偶蹄目,與河馬為姐妹群,因此未包含鯨類的偶蹄目是並系群(即部分演化支被歸入其他類群)。學界基於此修訂了原有的分類系統,將偶蹄目和鯨目合併為單系的 鯨偶蹄目(Cetartiodactyla),但在描述非鯨類的偶蹄動物時,慣用的稱法依然是偶蹄目(Artiodactyla)。
鯨與河馬的最近共同祖先,是5900萬年前由反芻動物分化出的半水生雜食動物,其中一支在5400萬年前的始新世從南亞等地入水生活,逐漸演化為完全水生的鯨類,並擴散至全球各大海域,部分種類還進入內陸河流及湖泊。在物種進化史上,一切脊椎動物均由3億年前海洋中的魚類演化而來,鯨類祖先在離開海洋2.5億年後重返海洋,被稱作“二次入水”,是進化史上一次罕見的重大轉折事件。
河馬的起源尚不明確,已知最早的化石是在肯亞發現的肯亞河馬屬(Kenyapotamus),生活在1600-800萬年前的中新世,相隔鯨類的分化有3000萬年的化石空白期。始新世中期出現的石炭獸科與河馬有著十分密切的聯繫,但是否為河馬的祖先仍未得到公認。
胼足亞目曾被視作反芻亞目的近親,因為它們體形相似,且都有反芻習性(儘管消化系統不同),但分子學的研究顯示胼足亞目更接近偶蹄目進化樹的根部,是現生偶蹄動物中最早分化的一批。胼足亞目曾非常多樣化,無防獸科、岳齒獸科、劍齒獸科等史前類群都被歸於此,唯一繁衍至今的是駱駝科,約在4620萬年前始新世中期發源於北美,上新世擴散到亞洲、北非和南美,現存3屬7種,其中4種為家畜。
豬形亞目現存兩支:舊大陸的豬科與新大陸的西猯科。該亞目曾被分為古齒獸下目、彎齒獸下目和 豬下目,其中彎齒獸下目是基於河馬建立的,收納了河馬科與史前的石炭獸科等,然而DNA測序表明河馬及其古代親系更接近鯨類和反芻亞目,因此3個下目的分類是無效的。豬形亞目的典型特徵是軀幹和四肢粗短,頭大且鼻吻部較長,適合發掘地下的食物;尾短小;體毛短而硬,頭頂至頸背部、雙耳及尾端具鬃毛;犬齒髮達且終身生長,成年雄性的上犬齒往往會突出嘴外,是打鬥時的利器。常在泥水中打滾以避免寄生蟲的侵擾,同時緩解炎熱。
反芻亞目擁有相當複雜的消化系統,是進化程度最高的陸生偶蹄目,也是偶蹄目中最繁盛的一類,現存6科:鼷鹿科、長頸鹿科、叉角羚科、鹿科、麝科、牛科(洞角科),其中牛科是偶蹄目中最大的科。這些科被劃歸2個下目:鼷鹿科是反芻亞目中最基底的一支,隸屬 鼷鹿下目;其他5科均屬 有角下目,5科中除麝科外,頭上均有雙角,但鹿科僅雄性有角(唯一的例外是馴鹿,兩性都有角),部分牛科也只有雄性有角。
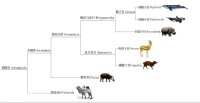
偶蹄目內部的分化關係
註:列表自偶蹄目往下逐級顯示,具體到科級,科級位置未定者,具體到屬級。列表僅收錄狹義的偶蹄類,未顯示鯨豚類(鯨下目)的分類情況。符號“†”表示已滅絕,“?”表示有疑問。
| 偶蹄目Artiodactyla(或鯨偶蹄目Cetartiodactyla) | ||
†Antiacodontidae †河豬科Choeropotamidae †麗豬科Leptochoeridae | ||
| 1 胼足亞目Tylopoda | ||
?†豬形獸屬Gobiohyus †Homacodontidae | ||
| †無防獸總科Anoplotherioidea | †無防獸科Anoplotheriidae †新獸科Cainotheriidae †Dacrytheriidae | |
| 駱駝總科Cameloidea | †鹿駝科Oromerycidae 駱駝科Camelidae | |
| †岳齒獸總科Merycoidodontoidea | †郊豬科Agriochoeridae †岳齒獸科Merycoidodontidae | |
| †劍齒獸總科Xiphodontoidea | †劍齒獸科Xiphodontidae | |
| 2全撰類Artiofabula | ||
| 2.1 豬形亞目Suina | ||
豬科Suidae 西猯科Tayassuidae †沙尼獸科Sanitheriidae | ||
| 2.2鯨反芻類Cetruminantia | ||
| 2.2.1反芻形類Ruminantiamorpha | ||
| 2.2.1.1 反芻亞目Ruminantia | ||
| 鼷鹿下目Tragulina | †疑芻駝科Amphimerycidae †Prodremotheriidae †原角鹿科Protoceratidae †異鼷鹿科Hypertragulidae †Praetragulidae 鼷鹿科Tragulidae †Archaeomerycidae †Lophiomerycidae | |
| 有角下目Pecora | †吉洛鹿科Gelocidae †Hoplitomerycidae †美鹿科Leptomerycidae | |
| 長頸鹿總科Giraffoidea | 長頸鹿科Giraffidae 叉角羚科Antilocapridae †梯角鹿科Climacoceratidae | |
| 鹿總科Cervoidea | †古鹿科Palaeomerycidae 鹿科Cervidae 麝科Moschidae | |
| 牛總科Bovoidea | 牛科Bovidae | |
| 2.2.2鯨凹齒形類Cetancodontamorpha | ||
?†安氏獸屬Andrewsarchus †豨科Entelodontidae | ||
| 2.2.2.1鯨河馬亞目Whippomorpha | ||
| 鯨形類Cetaceomorpha | †勞氏獸科Raoellidae | |
| 鯨下目Cetacea(此處省略其下級分類) | ||
| †雙錐齒獸總科Dichobunoidea | †雙錐齒獸科Dichobunidae †沼豬科Helohyidae †長尾豬科Cebochoeridae †混獸科Mixtotheriidae | |
| 凹齒下目Ancodonta | 河馬總科Hippopotamoidea | †石炭獸科Anthracotheriidae 河馬科Hippopotamidae |
現代陸地偶蹄目的第一趾完全退化,僅部分滅絕種類生有第一趾;四足的中軸通過第三趾和第四趾之間,此二趾同等發育,以對稱的形態均衡地承擔體重,是最發達的兩個足趾;第二趾和第五趾因種類而異,河馬第二、五趾的形態和功能齊全,與第三、四趾共同朝向前方支撐身體,而其他陸生偶蹄類的第二、五趾則相對細小,且懸於足後方不接觸地面,並無實際功能,甚至和第一趾一樣完全退化 。每個足趾前端都覆有鞘狀蹄甲,具保護作用,豬形亞目和大部分反芻亞目以趾端的蹄甲著地。
 偶蹄目 |  偶蹄目 |  偶蹄目 |
| 河馬每足四趾,功能健全 | 豬和大部分反芻亞目第二、五趾懸於足后,以第三、四趾的蹄甲著地 | 駱駝和小部分反芻亞目第二、五趾退化,趾甲未完全特化成蹄 |
偶蹄目胸腰部椎骨較奇蹄目為少,股骨無“第三轉子” 。門齒僅下頜門齒髮達,上門齒小或退化以至消失,僅以硬皮上唇咬合取食。前臼齒和臼齒在構造上均屬高冠齒,具有適於研磨的咀嚼面。例如豬的頰齒具有隆起呈多數的小瘤狀突,稱為丘齒型;反芻類的牛、羊和鹿的頰齒具有前後擴展V字形或新月形的牙齒。
有角下目動物的頭上有雙角,角大略可分為4種類型:1. 鹿科的角具有分枝,夏初長出的茸角覆以皮膚,后經骨質化,表皮脫離、光裸,繁殖期過後,鹿角在翌年春天脫落;2. 長頸鹿的角極短小,外覆有皮膚,但角和皮從不脫落,終生存在;3. 叉角羚的角不分枝,但有小叉,角鞘每年脫落;4. 牛科的角不脫落,不分叉,外套有角質硬鞘,角的骨心和角鞘終身生長,老齡動物角基粗,角干長。
一些豬類的腋窩到腹股溝部位有兩排乳房,而其他大部分偶蹄目只有一對或兩對乳房。
偶蹄目是哺乳綱中多樣性最高的目,最小的鼷鹿體重只有4千克,最大的藍鯨可達33米長、190噸重,為世界上最大的動物 。此外,河馬是偶蹄目中除鯨類外體型最大者,最重可超過3噸;長頸鹿則是現存最高的動物,頭頂距地面可超過5米。
關於鯨類的形態特徵,另見主詞條“鯨目”。
反芻亞目和胼足亞目的駱駝科具復胃,均有反芻特性。由於植物纖維較難消化,而採食過程一般比較匆忙,大部分食物未經充分咀嚼就吞咽下胃,因此反芻是為了縮短取食時間並加強消化粗糙食物而進化出的特殊功能。
反芻亞目的胃分四個腔室:瘤胃(內壁有許多瘤狀突起)、網胃(又稱蜂巢胃,內壁有形似蜂巢的網眼狀褶皺)、瓣胃(又稱重瓣胃,內壁有許多重疊而大小不等的書頁狀的葉瓣)、皺胃(又稱真胃,內壁多褶皺),前三個胃室無胃腺,總體作用是對食物進行發酵、過濾、磨碎以及營養成分的粗吸收,僅皺胃是分泌消化液的部分,相當於單胃動物的胃。瘤胃還有一個前庭,它有時也被看作是一個獨立的胃室,有些學者也將瘤胃與網胃合為一個單位,因此根據不同資料來源反芻亞目有四至五個胃室。駱駝科胃分三室,瓣胃和皺胃合而為一。豬形亞目和鯨河馬亞目的胃也有多個腔室(詳情因種類而異),但不反芻。
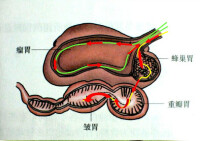
反芻亞目的胃(箭頭表示食物的流動方向)
偶蹄目中溫帶和寒帶地區的物種有固定的繁殖季節,熱帶地區的物種則全年繁殖。許多偶蹄目實行“一夫多妻制”,即雄性之間通過爭鬥決出單個優勝者,獲得一定範圍內與所有雌性的交配權。
偶蹄目的妊娠期普遍較長,產崽數較少。小型反芻類和豬類的妊娠期4-5個月不等;中型和大型鹿科、牛科及河馬為6-10個月;駱駝10-13個月;長頸鹿14-15個月。大部分豬類每胎產1-2個幼崽,小部分豬類每胎產崽可超過10個,是偶蹄目中產崽最多的。
偶蹄目幼體的發育程度極高,剛出生時便能睜開眼睛,且肢體功能完善,可在短時間內學會奔跑並融入群體。
多數偶蹄目的自然壽命為20-30年;和許多哺乳動物一樣,體型較大的種類壽命較長,反之較短。陸地偶蹄動物中,壽命最長的是河馬、母牛和駱駝,可達40-50年,而大型鯨類的壽命可超過100年。
偶蹄目動物對人類有極其重要的價值,是畜牧業和農業的重要組成部分,上萬年來都是人類最主要的肉、乳、毛皮製品來源,還被廣泛用於役使和娛樂活動。
