禽龍
鳥臀目禽龍科動物
徠禽龍(學名:Iguanodon,意為“鬣蜥的牙齒”),屬於蜥形綱鳥臀目鳥腳下目的禽龍類。禽龍是種大型鳥腳類恐龍,身長約9到10米,高4到5米,前手拇指有一尖爪,可能用來抵抗掠食者。它們主要生活在侏羅紀和白堊紀(距今6500萬—2億年)的大型恐龍。後肢發達,直立高約5米,從頭部至尾尖長11米。尾巴粗重,起平衡作用。素食,舌長,利牙鋸齒狀,用以撕扯和切碎樹葉。

禽龍骨架
身長約10米,高3到4米,前手拇指有一尖爪,可能用來抵抗掠食動物,或是協助進食。

禽龍
化位置大約位於行動敏捷的棱齒龍類首次出現,演化至鳥腳下目中最繁盛的鴨嘴龍類,這段過程的中間位置。此外,過去有許多化石被歸類於禽龍,年代橫跨侏羅紀啟莫里階到白堊紀森諾曼階,範圍廣達歐洲、北美洲、亞洲內蒙古、以及北非。但這些化石多被歸類於其他屬,或是建立為新屬。
對於禽龍的了解,因為新發現的化石而隨者時間不斷改變。禽龍的大量標本,包括從兩個著名屍骨層發現的接近完整骨骸,使得研究人員可提出許多禽龍生活方面的假設,包括進食、移動方式、以及社會行為。禽龍的重建圖也隨著標本的新發現而改變。
四足禽龍想像圖禽龍的手臂長(貝尼薩爾禽龍的前肢大約是後肢的75%長度)而粗壯,而手部相當不易彎曲,所以中間三個手指可以承受重量。拇指是圓錐尖狀,與中間三跟主要的指骨垂直。在早期重建圖裡,尖狀拇指被放置在禽龍的鼻子上。稍晚的化石則透露出拇指尖爪的正確位置,但它們的真實作用仍處於爭論中。它們可能用於防禦、或者搜索食物。小指呈修長、敏捷的,可能用來操作物體。後腿強壯,但並非用來奔跑,每個腳掌有三個腳趾。骨幹與尾巴由骨化肌腱支撐、堅挺(這些棒狀骨頭經常在模型或繪畫中省略)。禽龍與較晚期的近親鴨嘴龍類,在身體結構上相異不大。
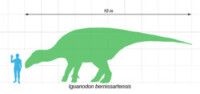
貝尼薩爾禽龍與人類的體型比較
禽龍 嘴前端較寬,長有骨質硬喙,長長的頜中後端密密麻麻排列著利齒,這一切都是為了能飽食到大量植物進化而來的結果。它的前肢較長,生有4根“手指”和一根像釘狀一樣尖的“拇指”,後肢強壯有力,並生有3個腳趾。但跑步時用兩條後肢,走路時一般都用四肢。它習慣用靈巧的“手”去抓扯植物,然後送入口中細嚼慢咽,悠悠品味,最後咽入胃中消化吸收。
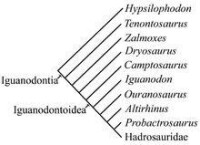
簡易版禽龍類演化樹
禽龍是古爬行動物,也是最早發現的恐龍。化石在1822年於英國南部發現。體長約9米,重7噸。前肢有五指,拇指像釘子。兩足行走,以植物為食。生存於白堊紀早期。化石主要發現於歐洲、北美、北非和東亞。

禽龍牙齒與現代鬣蜥的牙齒圖解
1840年在英格蘭梅德斯通發現的禽龍化石在確認過這些牙齒與鬣蜥牙齒的相似處后,曼特爾將它們命名為禽龍(Iguanodon);在希臘文里,iguana意為“鬣蜥”,odontos意為“牙齒”。曼特爾基於異速成長理論,而估計這動物的身長接近12米。
曼特爾最初是想將它命名為Iguanasaurus(鬣蜥龍),但他的朋友威廉·丹尼爾·科尼比爾(WilliamDanielConybeare)建議這個名字比較適合鬣蜥本身,而Iguanoides(似鬣蜥)或Iguanodon(鬣蜥牙齒)是更好的選擇。曼特爾當時忘記取種名,所以在1829年,弗里德里希·霍爾(FriedrichHoll)將禽龍命名為I.anglicum,後來修改為安格理克斯禽龍(I.anglicus)。

1834年在英格蘭梅德斯通發現的禽龍化石
同時,曼特爾與理查德·歐文(RichardOwen)之間產生了緊張關係,歐文是個著名科學家,有許多更好的發現,並且在英國混亂的改革時代中跟政治界與科學界有良好的社會關係。歐文是個堅定的創造論者,他反對當時引起爭論的早期演化理論,並將恐龍作為反擊的武器。在一項對於恐龍的研究中,歐文將恐龍的長度估計為超過61米,這使得恐龍並非只是巨型蜥蜴,並推演出它們是先進、類似哺乳類的動物;根據那個時代的理解,這些特徵是上帝所賦予的,而非從蜥蜴轉變為類似哺乳類的動物。
在曼特爾死前不久的1852年,他了解到禽龍並非重型、類似厚皮動物的動物,也就是說禽龍的外型並不如同歐文所認為的;但曼特爾的去世讓他無法參加水晶宮恐龍雕塑的創造,所以歐文版本的恐龍成為大眾所熟知的版本長達數十年。本傑明·沃特豪斯·霍金斯(BenjaminWaterhouseHawkins)有接近24個不同史前動物的重建雕塑,這些混凝土雕塑由鋼與磚瓦做成骨架;其中有兩個禽龍雕塑,一個呈四肢站立姿勢,另一個以腹部躺著。在四肢站立姿勢的禽龍完成之前,霍金斯曾邀請20個客人在骨架中舉辦宴會。
最大型的禽龍化石是在1878年發現於比利時貝尼沙特的煤礦坑,位在地表322米之下(1056呎)。在煤礦的管理者阿方斯·布里亞爾(AlphonseBriart)促進之下,路易·多洛(LouisDollo)與路易·德波夫(LouisdePauw)監視這些骨骸的出土,並重建它們。當地至少有38個禽龍個體被挖掘出土,其中大部分為成年個體。從1882年開始,這群化石中大部分被公開展覽;其中11個是以站立姿勢展出,而另20個是以它們被挖掘出土的型態展出。這些禽龍化石是布魯塞爾比利時皇家自然科學博物館的重要展覽品。其中一個的複製品在牛津大學自然歷史博物館展出。這32個化石中,大部分被歸類於新種貝尼薩爾禽龍(I.bernissartensis),比英國所發現的禽龍還要大型、粗壯;但其中一個化石,被歸類於不明確、纖細的曼氏禽龍(I.mantelli)。這些禽龍化石是已知第一群完整恐龍化石之一。除了這些禽龍之外,另外還發現了植物、魚類、以及其他爬行動物的化石,包含鱷類的伯尼斯鱷。
架設中的貝尼沙特禽龍骨骸在當時,化石保存技術才剛起步。而且,骨頭中的黃鐵礦會變化成硫酸鐵,破壞這些化石,使它們破碎、粉碎。當位在地表下時,化石保存於濕氣中,可避免變化發生;但一旦接觸到乾燥的空氣時,化學轉化便開始發生。那些位在布魯塞爾博物館的工作人員面對這個問題時,可能使用結合酒精、砒霜、以及蟲膠的物質來處理。這個綜合物質將同時滲透入化石(酒精),消滅所有生物媒介(砒霜),並使化石堅固(蟲膠)。這些綜合物質擁有意外的功效,可將濕氣保存在化石中,並延長破壞年限。現代的處理方式已不採用監視化石儲藏室的濕度,而是用聚乙二醇包復化石,並在真空幫浦中加熱,而濕氣將立即地被移除,而細小空間被聚乙二醇填滿,可密封並補強化石。
多洛利用這些貝尼沙特標本證明歐文將禽龍形容為厚皮動物,是不正確的。他將這些骨骸以鴯鶓與沙袋鼠的二足姿勢架設起來,並將原本放在鼻部的尖刺,重新置於拇指上。然而,多洛並非完全正確,但他當時是面對第一群完整恐龍化石,具有資訊與經驗上的劣勢。多洛的最大問題是他將禽龍尾巴彎曲。禽龍的尾巴因為硬化肌腱的原因,實際上將近筆直,如同這些化石剛出土的狀態。如果禽龍的尾巴以類似沙袋鼠或袋鼠的姿勢彎曲,它們的尾巴將會斷裂。如果禽龍的背部與尾巴是筆直狀態,當禽龍行走時,它們的身體將會與地面平行,而手臂則處於隨時支撐身體的狀態。
貝尼沙特地區的挖掘活動在1881年停止,但當地的化石並沒完全挖出土,仍有發現挖掘活動。在第一次世界大戰期間,當地被德意志帝國所佔據,德國古生物學家奧托·耶克爾(OttoJaekel)來到比利時,準備監督貝尼沙特的重新挖掘活動。在第一個含有化石的地層即將出土時,協約國重新佔領了貝尼沙特。該地區的挖掘活動因為財務問題而被阻礙了,並因為1921年的淹水而停止。
禽龍是種大型草食性動物,身長約10米,高3到4米,前手拇指有一尖爪,可能用來抵抗掠食動物,或是協助進食。
禽龍的化石多數發現於歐洲的比利時、英國、德國,它們主要生存於白堊紀早期的巴列姆階到早阿普第階,約1億3000萬年前到1億2000萬年前。演化位置大約位於行動敏捷的棱齒龍類首次出現,演化至鳥腳下目中最繁盛的鴨嘴龍類,這段過程的中間位置。此外,過去有許多化石被歸類於禽龍,年代橫跨侏羅紀啟莫里階到白堊紀森諾曼階,範圍廣達歐洲、北美洲、亞洲內蒙古、以及北非。但這些化石多被歸類於其他屬,或是建立為新屬。
並在1825年由英國地理學家吉迪恩·曼特爾進行描述與命名。斑龍、禽龍、以及林龍是用來定義恐龍總目的三個屬。禽龍與鴨嘴龍科共同屬於禽龍類演化支。
對於禽龍的了解,因為新發現的化石而隨者時間不斷改變。禽龍的大量標本,包括從兩個著名屍骨層發現的接近完整骨骸,使得研究人員可提出許多禽龍生活方面的假設,包括進食、移動方式、以及社會行為。禽龍的重建圖也隨著標本的新發現而改變。
貝尼薩爾禽龍與人類的體型比較
禽龍屬(Iguanodon)大型素食恐龍的統稱。化石見於歐洲、北非、亞洲東部廣大地區的上侏羅統和下白堊統。身長10公尺(米)多,頭部離地面4公尺(米)。這種兩足行走的動物的後肢很發達,長而粗的尾起平衡作用。前肢也較發達,具異常的前掌,朝上生長硬如尖釘的拇指與掌的其餘部分成直角。牙有鋸齒狀刃口。該屬是最早被發現和研究的恐龍。已找到許多完整個體的化石,有些成群被發現,表明他們曾成群行走。有人提出它具有部分水生的習性,當受到威脅時,進入河或湖中避難。
禽龍的化石多數發現於歐洲的比利時、英國、德國,此外也有一些可能是禽龍的化石出土於北美洲、亞洲內蒙古、以及北非。
禽龍是繼斑龍之後,世界上第二種正式命名的恐龍。禽龍的化石在1822年首次發現,並在1825年由英國地理學家吉迪恩·曼特爾進行新種描述。禽龍、斑龍、以及林龍為最初用來定義恐龍總目的三個屬。禽龍與鴨嘴龍科共同屬於禽龍類演化支。
對於禽龍的了解,因為新發現的化石而隨者時間不斷改變。禽龍大量的標本,包括從兩個著名河床發現的接近完整骨骸,使得研究人員可提出許多禽龍生活方面的假設,包括進食、運動、以及社會行為。禽龍的重建圖也隨者標本的新發現而改變。

禽龍
禽龍的牙齒類似鬣蜥的牙齒,但較大。鴨嘴龍科擁有多排不斷替換的牙齒,而禽龍在同一時間只有一副準備替換用的牙齒。上顎骨左右兩側最多各有29顆牙齒,前上顎骨則沒有牙齒,齒骨左右兩側則各有25顆牙齒;上下顎牙齒數量不一致的原因,是因為下顎的牙齒較寬。因為這些牙齒位於顎部外側,以及其他的生理特徵,禽龍被認為具有某種頰囊,可能由肌肉所構成,可以將食物置於兩頰咀嚼,如同大部分其他鳥臀目恐龍。
當禽龍的嘴部閉合時,上下顎的頰齒表面會互相磨合,可磨碎中間的食物,形成類似哺乳類的咀嚼動作。因為禽龍的牙齒是不斷替換的,所以它們能夠終生以堅硬的植物為食。另外,禽龍上下顎的前端缺乏牙齒,形成鈍狀的邊緣,可能復蓋著角質,可以咬斷樹枝。禽龍的小指纖細而靈活,可協助勾取食物。
不清楚禽龍平常以何種植物為食。較大的種可能以離地面4.5米以內的樹葉為食,例如貝尼薩爾禽龍。大衛·諾曼認為禽龍可能以木賊、蘇鐵、以及針葉樹為食,一般認為,白堊紀開花植物的出現與禽龍類有關,導因於這些恐龍以低高度植被為食。根據假設,由於禽龍類以裸子植物為食,使得類似草的早期被子植物有空間成長,但還沒有證據可以證明。無論禽龍以何種植物為食,根據它們的體型與繁盛,它們應該佔據者體型中到大型草食性動物的生態位。禽龍在英格蘭與以下恐龍共同生存:小型掠食者極鱷龍、大型掠食者始暴龍、重爪龍、新獵龍、小型草食性恐龍棱齒龍與荒漠龍、禽龍科的曼特爾龍、甲龍類的多刺甲龍、以及蜥腳下目的畸形龍。
曼特爾根據英格蘭梅德斯通的化石所繪製的禽龍重建圖早期的禽龍化石很破碎,使得科學家們對於禽龍的步態產生不同的看法。禽龍最初被描述成鼻上有角的四足動物。隨者更多的化石被發現,曼特爾發現禽龍的前肢遠短於後肢。他的對手理查德·歐文,則認為禽龍是種具有柱狀四肢的矮胖動物。恐龍的第一個原始比例重建工作被託付給曼特爾,但他因為健康不佳的原因而拒絕了,這些禽龍雕塑後來以歐文的版本作為原型。
在貝尼沙特發現了大量禽龍化石后,科學家們發現禽龍是種二足動物。但禽龍在當時被塑造成筆直站立的步態,尾巴拖曳在地面上,充當三腳架的第三支點。

古得利奇版本的重建圖
隨者禽龍的年齡增長,以及體重的增加,它們將更常採取四足步態;幼年貝尼薩爾禽龍的手臂較成年體的短,約是後肢長度的60%,成年個體的前肢長度則為後肢的70%。根據禽龍類的足跡化石,以及禽龍的手部、手臂結構,可推論禽龍採取四足步態時,中間三根蹄狀手指可支撐重量。禽龍的後腳掌相當長,上有三根腳趾,它們會採取趾行動物的方式,使用手指與趾爪來行走。禽龍以二足奔跑的最高速度估計為每小時24公里,但它們無法使用四足步態快速的奔跑。
在英格蘭的早白堊紀地層發現許多大型三趾足跡,尤其是在威特島的維耳德,這些足跡化石當初很難以敘述、解釋。有些早期的研究人員認為它們與恐龍有關係。在1846年,愛德華·泰戈特(EdwardTagart)將這些足跡化石歸類於生跡分類單元中的禽龍痕迹屬;而塞繆爾·比克爾斯(SamuelBeckles)則在1854年發現這些足跡類似鳥類的足跡,但可能來自於恐龍。在1857年,一個年輕禽龍的後肢被發現,足部擁有三根腳趾,顯示這些足跡可能來自於禽龍。儘管缺乏直接證據,這些足跡化石常被歸類於禽龍。在英格蘭的一個足跡化石顯示禽龍可能以四足方式行進,但足跡本身保存狀態不佳,因此很難作為直接證據。被歸類於禽龍痕迹屬的足跡化石,位於歐洲挪威的卑爾根群島與司瓦爾巴特群島,皆為發現禽龍化石的地方。
最早發現的禽龍拇趾尖爪,在1840年發現於德國美斯頓拇指的尖爪是禽龍的最著名特徵之一。雖然曼特爾最初將拇指尖爪放置在禽龍的鼻部上,但道羅根據在貝尼沙特發現的完整標本,將拇指尖爪放置於手部的正確位置上。但自從80年代以來,仍有許多恐龍的大型拇指尖爪被錯置在足部,類似馳龍科,例如西北阿根廷龍、拜倫龍、以及大盜龍。
禽龍的拇指尖爪被認為是種對付掠食者的近身武器,類似短劍,但也可能用來挖開水果與種子,甚至用來與其它禽龍打鬥。一位科學家認為這個拇指尖爪連接者毒腺,但這觀點並沒有被接受,因為尖爪的內部並非中空,表層也沒有溝槽可使毒液流動。
貝尼沙特的大量禽龍化石,有時被認為是單一的災害而造成的,還有被認為是多種原因而造成的。根據群體生活的說法,該地至少保存了三種死亡方式,而大量的個體在很短的地質時間內死亡(約10到100年內),但這並不代表禽龍是種群居動物。貝尼沙特的幼年禽龍化石非常普遍,不同於現代群體動物的死亡模式。可能是周期性的洪水將大量的屍體沖積到湖泊或沼澤。
德國Nehden鎮的禽龍化石,則具有較大的年齡範圍,甚至是曼特爾龍與貝尼薩爾禽龍的六倍;根據地理特性,顯示可能有群居動物曾遷徙經過河邊。
不像其他被假設的群居動物,例如鴨嘴龍類與角龍科,沒有證據顯示禽龍為兩性異形動物。有論點認為貝尼沙特的曼氏禽龍或阿瑟菲爾德禽龍(目前都為曼特爾龍)代表禽龍的某種性別,可能是雌性,而較大、較結實的貝尼薩爾禽龍,則可能為雄性。然而,這個理論沒有得到任何支持。
禽龍屬

禽龍
最初的模式種是安格理克斯禽龍(I.anglicus),但該種的模式標本只有一個牙齒,而且該種自從被發現以來,只有一個部分化石。因此在2000年3月,國際動物命名法委員會將禽龍的模式種改為較著名的貝尼薩爾禽龍。禽龍的最初牙齒目前位在紐西蘭威靈頓的紐西蘭國家博物館,但並沒有被展出。禽龍的最初牙齒原本為吉迪恩·曼特爾(GideonMantell)所有,當曼特爾死後,他的化石被交給住在紐西蘭的兒子沃爾特·曼特爾(WalterMantell)。
禽龍雕塑,位於布魯塞爾比利時皇家自然科學研究院外部在眾多被歸類於禽龍屬的種中,只有少數被認為是有效種。
貝尼薩爾禽龍(I.bernissartensis):禽龍的模式種,由喬治·亞伯特·布蘭潔(GeorgeAlbertBoulenger)在1881年命名。貝尼薩爾禽龍是因為貝尼沙特所發現的眾多骨骸而著名,也因為在歐洲各地的發現而著名。貝尼薩爾禽龍也包含了在蒙古發現的東方禽龍(I.orientalis),東方禽龍的目前狀態為疑名。
道氏禽龍(I.dawsoni):是由理查德·萊德克(RichardLydekker)在1889年命名。是萊德克在1880年代晚期命名的兩個種之一,這兩個種皆為有效種,但很少受到討論。道氏禽龍的化石包含兩個部分骨骸,發現於英格蘭東薩塞克斯郡的喜士定地層,年代為下白堊紀凡藍今階與巴列姆階。
菲頓禽龍(I.fittoni):也由萊德克在1888年命名,也是在英格蘭東薩塞克斯郡的Hastings地層發現,另外在西班牙也有發現,已發現三個部分骨骸。道氏禽龍與菲頓禽龍的差異在於脊椎與骨盆的特徵、形狀,以及體格大小。道氏禽龍的體格較菲頓禽龍健壯,擁有類似彎龍的大型脊椎骨,上有短神經棘;而菲頓禽龍的神經棘長而狹窄,角度陡峭。
拉科塔禽龍(I.lakotaensis):是由大衛·威顯穆沛與菲利浦·比約克(PhilipR.Bjork)在1989年所命名。拉科塔禽龍是北美洲發現的唯一禽龍有效種,化石包含一個部分頭顱骨,發現於美國南達科他州的拉科塔組,年代為下白堊紀的巴列姆階。拉科塔禽龍的歸類是有爭議的,有些研究人員認為它們比貝尼薩爾禽龍原始,並與Theiophytalia有接近親緣關係,而大衛·諾曼(DavidNorman)則認為它們是貝尼薩爾禽龍的一個異名。

禽龍標本
1874年發現於多塞特郡的波白克層,年代為上侏儸紀提通階到下白堊紀巴列姆階,已經被大衛·諾曼與保羅·巴雷特(PaulBarrett)重新歸類於彎龍;大禽龍(I.major)的化石包含一個發現於威特島的脊椎骨,由歐文在1842年敘述為扭椎龍的一個種,狀態為疑名,現在則被認為是安格里可斯禽龍的一個異名。
除了歐文所命名的這兩個種以外,還有13個種被重新歸類於其他屬。
阿爾比斯禽龍(Iguanodonalbinus):是由捷克古生物學家安東寧·弗里奇(AntoninFritsch)在1893年命名,被懷疑是非恐龍的爬行動物,目前為阿爾比斯龍。
阿瑟菲爾德禽龍(I.atherfieldensis):由雷金納德·胡利(ReginaldHooley)在1925年命名,比貝尼薩爾禽龍較小、較不粗壯,而有較長的神經棘,在2006年重新命名為曼特爾龍。
I.exogyrarum:由弗里奇在1878年命名,有時也拼成I.exogirarum或I.exogirarus。化石材料非常少,因此狀態為疑名,目前被喬治·奧利舍夫斯基(GeorgeOlshevsky)重新歸類於Ponerosteus。
凡登禽龍(I.valdensis):是由約翰·赫克(JoneHulke)在1879年命名,化石包含脊椎與骨盆,發現於威特島的巴列姆階;最初被命名為威特島龍,可能是曼特爾龍的一個成長中個體。
纖細禽龍(I.gracilis):是由萊德克在1888年命名,當時為楔椎龍(Sphenospondylus)的模式種,在1969年被洛尼·史提爾(RodneySteel)歸類於禽龍;可能是曼特爾龍。
福氏禽龍(I.foxii):最初是在1869年由湯瑪斯·亨利·赫胥黎(ThomasHenryHuxley)命名為棱齒龍的模式種,歐文在1873年或1874年將它重新歸類於禽龍,但不久后被回復成棱齒龍。
荷林頓禽龍(I.hollingtoniensis):由萊德克在1889年所命名,是菲頓禽龍的異名。
普萊斯特維奇禽龍(I.prestwichii):是由約翰·赫克在1880年命名,已被重新歸類於彎龍。
絲利禽龍(I.seeleyi):由約翰·赫克在1882年命名,被認為與貝尼薩爾禽龍是同一種動物。
休斯禽龍(I.suessii):由埃曼努埃爾·邦澤爾(EmanuelBunzel)在1871年命名,后被重新歸類於柵齒龍。
曼氏禽龍(I.mantelli):由汪邁爾在1832年命名,但汪邁爾當初所研究的化石,與用來命名安格理克斯禽龍的化石相同;然而,數個梅特斯通的標本,以及一個貝尼沙特的化石,長期以來被歸類於曼氏禽龍。較纖細的貝尼沙特化石,已被重新歸類於曼特爾龍。
東方禽龍(I.orientalis):是由蘇聯古生物學家阿納托利·康斯坦丁諾維奇·羅特傑斯特文斯基(AnatolyKonstantinovichRozhdestvensky)在1952年命名,化石材料不多。其中一個頭顱骨具有明顯拱起的口鼻部,因此該頭顱骨在1998年被重新命名為高吻龍。而東方禽龍因為無法與貝尼薩爾禽龍區別開來,因此目前的狀態為疑名。
菲利浦禽龍(I.phillipsi):由哈利·絲利(HarrySeeley)在1869年命名,后被重新歸類於頜鋸齒龍。
目前有5個禽龍物種是疑名或未經描述。
安格理克斯禽龍(I.anglicus):在1829年由弗里德里希·霍爾(FriedrichHoll)命名,是禽龍最初的模式種,後來被貝尼沙爾禽龍所取代。安格里可斯禽龍有時也被拼為I.angelicus與I.anglicum。該種的化石只有牙齒,發現於英格蘭的東薩塞克斯郡,年代為下白堊紀的凡蘭吟階與巴列姆階。
希氏禽龍(I.hillii):由埃德溫·塔利·牛頓(EdwinTullyNewton)在1892年發現,化石只有一顆牙齒,發現於赫福郡的下白堊層,年代為上白堊紀早森諾曼階,是某種早期鴨嘴龍科恐龍。
徠蒙古禽龍(I.mongolensis):狀態為疑名,是根據書籍上的照片來命名,化石後來被命名為高吻龍。
奧廷格禽龍(I.ottingeri):由彼得·高爾頓(PeterGalton)與詹姆斯·A·詹森(JamesA.Jensen)在1979年命名,化石只有牙齒,發現於猶他州的下西德山組,年代可能為阿普第階,狀態也為疑名。
先驅禽龍(I.praecursor):由E·紹瓦熱(E.Sauvage)在1876年命名,化石只有牙齒,發現於法國加萊海峽省,年代為晚侏儸紀啟莫里階,其實是種蜥腳下目恐龍,有時被歸類於新牙龍,但先驅禽龍與新牙龍來自於不同的地層組。
其他所知有限的屬與種,被認為於禽龍的不確定種,相較於重新命名的阿瑟菲爾德禽龍,它們的歸類更不確定。這些種包含:Heterosaurusneocomiensis、Hikanodon、Therosaurus、扭椎龍的一個種(Streptospondylusrecentior)、短尾鯨龍(Cetiosaurusbrachyurus)、以及一部分的矮鯨龍(C.brevis)化石(嵌合體)。原禽龍是個無資格名稱(Nomennudum),可能是扭椎龍中非常不確定的大扭椎龍(S.grandis)與邁氏扭椎龍(S.meyeri)兩個種。

基本信息
- 中文名
- 禽龍
- 外文名
- Iguana tooth
- 別名
- 鬣蜥的牙齒
- 拉丁學名
- Iguanodon
- 界
- 動物界
- 亞門
- 脊椎動物亞門
- 門
- 脊索動物門
- 綱
- 爬行綱
- 亞綱
- 雙孔亞綱
- 目
- 鳥臀目
- 亞目
- 角足亞目、鳥腳亞目
- 科
- 禽龍科
- 屬
- 禽龍屬
- 分布區域
- 歐洲、北美、北非和東亞
- 命名者及年代
- 白堊紀早期