細胞膜受體
細胞膜受體
細胞膜受體(cell membrane receptor)是細胞表面的一種或一類分子,它們能識別、結合專一的生物活性物質(稱配體),生成的複合物能激活和啟動一系列物理化學變化,從而導致該物質的最終生物效應。細胞環境中各種因素的變化,是通過細胞膜受體的作用而影響細胞內的生理過程發生相應的變化。
細胞膜受體也是鑲嵌在膜脂質雙分子層中的膜蛋白質。受體蛋白質一般由兩個亞單位組成:裸露於細胞膜外表面的部分叫調節亞單位,即一般所說的受體,它能“識別”環境中的特異化學物質(如激素、神經遞質、抗原、藥物等)並與之結合;裸露於細胞內表面的部份叫催化亞單位,常見的是無活性的腺苷酸環化酶(AC)。一般將能被受體識別的環境中的特異化學物質叫信號或叫配體。配體所作用的細胞又叫那個配體的靶細胞。一般來講,受體與配體的結合有高度特異性。當某一配體與其靶細胞膜上的特異性受體結合時,調節亞單位構型變化,隨即激活膜上的AC,在Mg2+存在的條件下,AC催化細胞內一系列生物化學反應,進而引起靶細胞生理功能的改變。也有的膜受體與配體結合后並不繼發細胞內一系列生化反應,而是通過改變細胞膜對離子的通透性而產生生理效應。
“受體”這個名詞在19世紀末20世紀初已經開始應用,當時認為藥物作用時必須與靶細胞上與之互補的分子——受體——相結合后才能發揮效應。後來受體概念的應用逐漸擴展到藥物以外的其他信息分子作用於細胞的過程。
由於細胞內受體含量極微,有些受體穩定性又差,因此受體的分離、純化比較困難。迄今只有從電鰩和電鰻的電器官中分離的乙醯膽鹼的煙鹼膽鹼能受體和從正常人胎盤中分離的胰島素受體已經得到純度很高、數量足夠的樣品,因而對它們的結構也有了較多的了解。電鰩的乙醯膽鹼的煙鹼膽鹼能受體是由分子量為26000~64000的四種亞基組成,從電鰻分離的受體則由分子量為42000和54000二種亞基組成。電子顯微鏡觀察的結果表明,這種受體由5~6個直徑為3~4納米的亞基構成玫瑰花結狀結構,其直徑約8~9納米,中心可能是離子通道。胰島素受體的斯托克半徑約68~72埃,分子量約30~100萬,它主要由3種亞基:α(分子量約135000),β(分子量約95000)和γ(分子量約45000)組成。α亞基為結合亞基,亞基之間靠—S—S—相連,亞基內部還有—S—S一結構。
一種細胞膜可以含有幾種不同的受體,如脂肪細胞膜上含有腎上腺素、胰高血糖素、胰島素等近10種激素受體。它們的數目互不相同。同一受體在不同細胞膜上的受體數目也是不同的。一般的受體的密度為103~104個/細胞,但電鰩電器官上的乙醯膽鹼的受體的密度和數量較大,分別為104~105微米2或1011個/細胞。受體的數目在正常生理條件下是恆定的,但由於細胞生理狀態不同(如生長速度,分化程度,細胞周期等)和外界環境變化的影響,也會發生一定的改變。
受體在膜上的分佈是不均勻的,經常相互聚集形成“簇”的形式。
受體調節的因素和途徑很複雜,在正常生理情況下受體數目受微環境影響而上升或下降,稱為上升或下降調節。其中與受體結合的配體濃度對調節受體具有較重要的作用,例如,當動物或人的血液中胰島素濃度較高時,靶細胞上的胰島素受體濃度即下降,如果胰島素濃度降低時,受體數目會迅速上升。受體的調節還可通過“負協同效應”來進行,即:受體-受體的相互作用會導致受體的親和力發生變化。例如,受體與胰島素分子的結合會促使受體與周圍其他受體之間的相互作用,導致受體的親和力下降,從而使胰島素分子從受體上解離下來。
在機體內已經發現某些受體的自身抗體,例如,1975年美國從一種β型嚴重胰島素抵抗症病人中發現有胰島素受體的自身抗體。這些抗體與受體結合可模擬胰島素的許多作用(例如,抑制脂肪分解,刺激葡萄糖的轉移和利用),但它會逐漸降低細胞對受體被結合后的生物化學反應的敏感性。加之抗體的存在也會降低受體對胰島素的親和力,結果在臨床上就表現為嚴重的胰島素抵抗症。
除胰島素受體外,兒茶酚胺受體,乙醯膽鹼的煙鹼膽鹼能受體,促甲狀腺素受體在病理情況下都發現有相應的自身抗體存在。
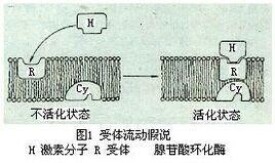
激素與受體結合后如何產生生物效應?20世紀60年代提出的第二信使假設認為,作為第一信使的激素分子與細胞膜受體結合后並不進入細胞。結合激素的受體能使位於膜上的腺苷酸環化酶活化,從而使ATP轉成環(化)腺苷酸(cAMP),後者稱為第二信使,它能引發細胞內一系列生化反應而產生最終生物效應。例如,腎上腺素在腎上腺髓質分泌后通過血液輸送至肝細胞產生效應。它們與肝細胞表面受體相結合后能使膜上腺苷酸環化酶活化催化ATP形成cAMP,後者使蛋白激酶等一系列酶蛋白相繼活化,最終使糖原分解成葡萄糖,從而使血糖濃度升高。其他一些激素,例如,促腎上腺皮質激素(ACTH),促卵泡激素(FSH),胰高血糖素和兒茶酚胺激素等在作用時也都是分別與它們的靶細胞受體相結合,導致腺苷酸環化酶的激活產生cAMP。這些激素在細胞膜上具有各自專一的受體,但腺苷酸環化酶卻是共用的。待激素分子與膜受體相結合形成複合體后,它們在膜上進行側向移動,一旦與腺苷酸環化酶相遇偶聯才能導致後者的構象變化而被激活。一旦激素稀釋或解離,受體和腺苷酸環化酶便又經移動而回到非偶聯狀態。這稱為“受體流動假說”(圖1)。
有些受體,如α類腎上腺素能受體與激素結合后細胞內不是cAMP而是Ca2+的濃度有明顯增加,這可能是由於膜上Ca2+通道開啟,使細胞外Ca2+進入細胞引起的。Ca2+能與鈣調蛋白結合併引起一系列反應,因而有人認為Ca2+也是一種第二信使。
近年來,發現有些激素,如胰島素,與細胞膜受體結合后並不僅僅停留在表面,形成的胰島素-受體複合物可通過內吞方式進入細胞內部(見內吞與外排),繼而這些複合物再與溶酶體融合或與其他細胞器膜相結合而產生生物效應。
利用高純度的胰島素研究表明,胰島素與受體的α亞基結合后可使受體本身固有的某種蛋白激酶活化,使受體分子本身,主要是β亞基發生磷酸化作用,這稱為自身磷酸化作用,它繼而又引起一連串的磷酸化反應,這可能是胰島素產生生物效應過程中一個十分重要的環節。
細胞膜受體
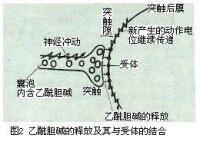
在中樞神經系統,前一神經原的軸突末梢興奮時,突觸處細胞質內大量儲存泡通過與突觸前膜相融合而將所含的乙醯膽鹼釋放至距離為0.5納米的突觸隙,之後乙醯膽鹼與突觸后膜上的受體相結合,使突觸后的神經原發生興奮,從而使神經衝動傳遞下去(圖2)。釋放的乙醯膽鹼如果作用於效應器(如肌肉),就能引起反應(如收縮)。當乙醯膽鹼與肌肉細胞受體結合后,會引起它的構象發生變化,打開離子通道,使膜對Na+、K+的通透性瞬時增加,從而導致膜電位的去極化,使肌肉收縮。一個與乙醯膽鹼結合了的受體可導致5×104離子通過,顯示了受體的放大作用。
現在發現很多毒素也是通過與細胞膜上的受體相結合后才產生效應的。如霍亂毒素是霍亂弧菌產生的外毒素,分子量為84000,由A、B二種亞單位組成。A亞單位有兩條肽鏈A1和A2,由一對二硫鍵聯接。亞單位B與細胞膜上的受體相結合。亞單位A1則具有激活膜上腺苷酸環化酶的作用。
霍亂毒素的受體是一種神經節苷脂,毒素與它結合后可能會發生一系列反應,如首先引發受體的構象變化,接著亞單位A1在激活腺苷酸環化酶的過程中將煙醯胺腺嘌呤二核苷酸(NAD)中的腺二磷核糖轉移到細胞膜中的一種蛋白質上。在正常情況下,鳥三磷通過與這種蛋白質相結合使腺苷酸環化酶激活,待鳥三磷被鳥三磷酶水解,激活作用即停止。但是如果鳥三磷結合在含有腺二磷核糖的蛋白質上就不易被水解,因而延長了腺苷酸環化酶作用的時間,產生的生物效應的持續時間也比較長。
霍亂毒素的作用與激素有幾點不同:①激素的啟動時間很短,霍亂毒素常常要延遲1~3刻鐘后才開始;②激素去除后,效應很快消失,而霍亂毒素在去除後生物效應還可持續幾小時到幾天;③一種激素的受體只在少數幾種細胞中存在,而霍亂毒素的受體在很多細胞膜上都有分佈。
凝集素是從各種植物或低等動物組織中分離而得到的一類特殊蛋白質,能與動植物細胞表面的受體發生特異性結合產生一系列的生理效應。它們通過與細胞表面寡糖結構決定簇的交互作用導致細胞發生凝集,因而又稱凝集素。最早發現它們能誘導紅細胞發生凝集而用於臨床血型分類,故又有植物血凝素之稱。目前已報道的凝集素多達500種。
與凝集素特異結合的受體位於細胞表面,具有複雜的寡糖鏈結構,它們擁有特殊的聯結、側鏈及多肽附著物。在這些寡糖鏈間還存在非共價鍵(如氫鍵)的相互作用。凝集素與受體寡糖鏈的結合部位以往都認為是末端的糖殘基,現在證明還能與多糖核心部位相結合。
不同的凝集素可以各有專一的受體,但也可以具有共同的受體。總的來講,凝集素受體的專一性比抗原-抗體的專一性要差。
凝集素使細胞凝集是由於一個凝集分子能與細胞表面兩個或兩個以上的結合點結合后所引起的。
當凝集素與其專一受體結合后可引發細胞多種生理效應,如細胞凝集,特別是腫瘤或轉化細胞可發生高效價的凝集作用;誘發淋巴細胞發生絲裂現象以及對膜透性、受精過程、吞噬及細胞增殖等都可產生一定的影響。
基本信息
- 中文名
- 細胞膜受體
- 外文名
- cell membrane receptor
- 作用
- 能識別、結合專一的生物活性物質
- 解釋
- 細胞表面的一種或一類分子