鞘磷脂
組成細胞膜的鞘脂
鞘磷脂存在於大多數哺乳動物細胞的質膜內,是髓鞘的主要成分。
鞘磷脂是在絲氨酸棕櫚醯轉移酶(serine palmi-toyltransferase, SPT)、3-酮 基 二 氫 鞘 氨 醇 還 原 酶(3-ketosphinganine reductase)、神經醯胺合成酶、二氫神經醯胺脫氫酶(dihydroceramide desaturase)和鞘磷脂合成酶(sphingomyelin synthase, SMS)的催化下進行生物合成的。鞘磷脂合成是神經醯胺轉化的主要途徑,是通過SMS催化的。
鞘磷脂是細胞膜的主要組成成分,其代謝產物如神經醯胺(ceramide, Cer)、鞘氨醇(sphingosine, Sph)、1-磷酸鞘氨醇(sphingosine-1-phosphate, S1P)是具有生物活性的信號分子,可作為第一和(或)第二信使調控細胞的生命活動,如細胞的生長、分化、衰老和凋亡等許多重要的信號轉導過程。
這些鞘磷脂的代謝產物參與細胞增殖與凋亡的調控:Cer和Sph是細胞增殖的負調控子,能夠抑制細胞生長、促進細胞凋亡;而其進一步的代謝物S1P則刺激細胞生長、抑制細胞凋亡。Cer/Sph和S1P構成了一個重要的代謝平衡體,它們之間的動態平衡與細胞的生存、死亡和生命活動密切相關。有人將由Cer、Sph和S1P共同構成的動態體系,稱作“鞘磷脂變阻器”(Sphingoliiic Rhecostat)。
鞘磷脂是動物細胞膜及其他各種生物膜的主要組成部分。鞘磷脂可以被鞘磷脂酶水解,生成神經醯胺等一系列生物活性脂。神經醯胺作為細胞第二信使,在心血管系統可以產生多樣的生物學效應:
(1)誘導包括平滑肌及心肌細胞在內的各種細胞凋亡;
(2)調節平滑肌細胞的分化和增殖;
(3)調節免疫功能和炎症反應;
(4)參與氧化應激。
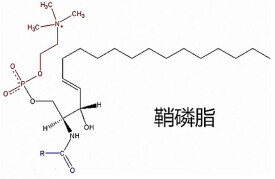
鞘磷脂極性頭部分是磷脂醯膽鹼或磷脂醯乙醇胺。
鞘磷脂結構與甘油磷脂相似,因此性質與甘油磷脂基本相同。
鞘磷脂是哺乳動物血漿中第二豐富的磷脂,可見於所有的主要脂蛋白中。高達18%的血漿磷脂以鞘磷脂的形式存在,在不同亞型的脂蛋白中,磷脂醯膽鹼與鞘磷脂的比值差異較大。致動脈粥樣硬化的脂蛋白,如VLDL和LDL都是富含鞘磷脂的。動脈粥樣硬化病變中鞘磷脂的含量高於正常動脈組織。
抑制鞘磷脂的從頭合成明顯抑制動脈粥樣硬化形成。其可能的機製為:
①減少致動脈粥樣硬化脂蛋白中的鞘磷脂含量可以減少其在主動脈的聚集;
基本信息
- 外文名
- sphingomyelin
- 結構
- 與甘油磷脂相似
- 性質
- 與甘油磷脂基本相同。