半保留複製
半保留複製
半保留複製(semiconservative replication):一種雙鏈脫氧核糖核酸(DNA)的複製模型,其中親代雙鏈分離后,每條單鏈均作為新鏈合成的模板。因此,複製完成時將有兩個子代DNA分子,每個分子的核苷酸序列均與親代分子相同。子代DNA分子中,一條鏈來自親代,另一條鏈為新合成的鏈。這是1953年沃森(J.D.Watson)和克里克(F.H.C.Crick)在DNA雙螺旋結構基礎上提出的假說,1958年得到實驗證實。
半保留複製闡述了在所有已知細胞中DNA複製的機制。半保留複製的名字來源於這樣的事實,在複製產生的兩個子代DNA拷貝中,每個拷貝的DNA雙鏈包含一個來自親代DNA的單鏈和一個新合成的DNA單鏈。
DNA的半保留複製假說最早由前蘇聯生物學家尼古拉·科爾佐夫(Nikolai Koltsov)於1927年提出。1953年沃森(J.D.Watson)和克里克(F.H.C.Crick)發表的DNA雙螺旋結構為此假說提供了結構上的依據。1958年美國科學家馬修 梅塞森(Matthew Meselson)和富蘭克林 斯塔爾(Franklin Stahl )的DNA同位素標記試驗 證實了DNA的雙螺旋結構和半保留複製機制。
DNA既然是主要的遺傳物質,它必須具備自我複製的能力,即通過複製形成新的和原來一樣的DNA分子的能力。但雙鏈DNA是如何解鏈、如何進行複製和如何保證DNA序列不變的,一直有很多的假說。
DNA在活體內的半保留複製特徵已為1958年以來的大量試驗所證實。DNA分子獨特的雙螺旋結構,為複製提供了精確的模板,通過鹼基互補配對,保證了複製能夠準確地進行。DNA分子通過複製,使遺傳信息從親代傳給了子代,從而保持了遺傳信息的連續性。DNA的這種複製方式對保持生物遺傳的穩定具有非常重要的作用。
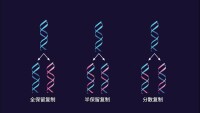
DNA還可能存在其他兩種複製方式,都以原來親本DNA雙鏈分子作為模板鏈。
全保留複製(conservative replication):保守複製會使兩條原始模板DNA鏈以雙螺旋結合在一起,併產生由兩條含有所有新DNA鹼基對的新鏈組成的拷貝。
分散複製(dispersive replication):分散複製將產生兩個DNA拷貝,兩個拷貝都含有由原始鏈或兩個新鏈組成的不同DNA區域。
半保留複製
DNA既然是主要的遺傳物質,它必須具備自我複製的能力。瓦特森和克里克(1953)在提出DNA雙螺旋結構模型的同時,對DNA複製也進行了假設。他們根據DNA分子雙螺旋結構模型,認為DNA分子的複製,首先是從它的一端氫鍵逐漸斷開。當雙螺旋的一端已拆開為兩條單鏈時,各自可以作為模板,從細胞核內吸取與自己鹼基互補的遊離核苷酸(A吸取T,C吸取G),進行氫鍵的結合,在複雜的酶系統的作用下,逐漸連接起來,各自形成一條新的互補鏈,與原來模板單鏈互相盤旋在一起,兩條分開的單鏈恢復為雙鏈DNA分子,與原來的完全一樣。DNA的這種複製方式稱為半保留複製(semiconservative replication),因為通過複製所形成的新的DNA分子,保留原來親本DNA雙鏈分子的一條單鏈。
DNA在活體內的半保留複製特徵已為1958年以來的大量試驗所證實。DNA的這種複製方式對保持生物遺傳的穩定具有非常重要的作用。
還可能存在其他兩種複製方式,都以原來親本DNA雙鏈分子作為模板鏈。一種方法稱為全保留複製(conservative replication),在複製過程中新的DNA分子單鏈結合在一起,形成一條新的DNA雙鏈,而親本DNA雙鏈仍然被保留在一起。另一種方法稱為散布式複製(dispersive replication),在複製過程中親本DNA雙鏈被切割成小片段,分散在新合成的兩條DNA雙鏈分子中。
半保留複製
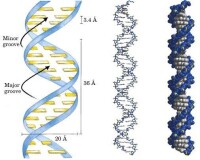
構成DNA分子的基本單位是脫氧核苷酸,許許多多脫氧核苷酸通過一定的化學鍵連接起來形成脫氧核苷酸鏈,每個DNA分子是由兩條脫氧核苷酸鏈組成。DNA分子結構的特點是:①DNA分子的基本骨架是磷酸和脫氧核糖交替排列的兩條主鏈;②兩條主鏈是平行但反向,盤旋成的規則的雙螺旋結構,一般是右手螺旋,排列於DNA分子的外側;③兩條鏈之間是通過鹼基配對連接在一起,鹼基與鹼基間是通過氫鍵配對在一起的,其中A與T以2個氫鍵相配對,C與G之間以3個氫鍵配對。所以在一個DNA分子中,G和C的比例較高,則該DNA分子就比較穩定。
DNA分子結構具有相對的穩定性是由兩個方面決定的。一是基本骨架部分的兩條長鏈是由磷酸和脫氧核糖相間排列的順序穩定不變;二是空間結構一般都是右旋的雙螺旋結構。DNA分子的穩定性主要由三種化學鍵維持:互補鹼基之間的氫鍵,鹼基之間的鹼基堆集力,以及主鏈上帶負電的磷酸與溶液陽離子之間的離子鍵。DNA 分子的多樣性是由鹼基對的排列順序的多樣性決定的。DNA分子的特異性是指對於控制某一特定性狀的DNA分子中的鹼基排列順序是穩定不變的,如控制合成唾液澱粉酶的基因中,不論是何人,這段DNA分子中的鹼基排列順序是穩定不變的。
半保留複製
由四種脫氧核苷酸組成的DNA分子為什麼能夠成為遺傳物質,仍然
感到困惑不解。為此,許多科學家都投入到對DNA分子結構的研究
中。1953年,美國科學家沃森和英國科學家克里克,共同提出了
DNA分子的雙螺旋結構模型(如圖)。
DNA分子的結構模式圖中可以看出,DNA分子的基本單位是脫氧核苷酸(如圖)。由於組成脫氧核苷酸的鹼基只有四種:腺嘌呤(A)、鳥嘌呤(G)、胞嘧啶(C)和胸腺嘧啶(T),因此,脫氧核苷酸也有四種,即腺嘌呤脫氧核苷酸、鳥嘌呤脫氧核苷酸、胞嘧啶脫氧核苷酸和胸腺嘧啶脫氧核苷酸。DNA分子就是由很多個脫氧核苷酸聚合而成的長鏈,簡稱多核苷酸鏈。
沃森和克里克認為,DNA分子的立體結構是規則的雙螺旋結構。這種結構的主要特點是:(1)DNA分子是由兩條鏈組成的,這兩條鏈按反向平行方式盤旋成雙螺旋結構。(2)DNA分子中的脫氧核糖和磷酸交替連結,排列在外側,構成基本骨架;鹼基排列在內側。(3)DNA分子兩條鏈上的鹼基通過氫鍵連結成鹼基對,並且鹼基配對有一定的規律:A(腺嘌呤)一定與T(胸腺嘧啶)配對;G(鳥嘌呤)一定與C(胞嘧啶)配對。鹼基之間的這種一一對應關係,叫做鹼基互補配對原則。在DNA分子的結構中,鹼基之間的氫鍵具有固定的數目,即A與T之間以2個化學鍵相連(A=T),G與C之間以3個化學鍵相連(G≡C)。由於嘌呤分子(A、G)大於嘧啶的分子(C、T),因此,要保持DNA兩條長鏈之間的距離不變,必定是一個嘌呤與一個嘧啶配對。根據鹼基分子所佔空間的大小,只有A與T配對,G與C配對,鹼基對的長度才能大致相同。根據DNA分子的上述特點,沃森和克里克製作出了DNA分子的雙螺旋結構模型。
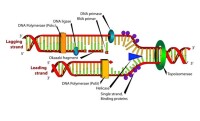
從製作的DNA雙螺旋結構模型中可以看出,組成DNA分子的鹼基雖然只有四種,但是,鹼基對的排列順序卻是可以千變萬化的。例如,在生物體內,一個最短的DNA分子也大約有4000個鹼基對,這些鹼基對可能的排列方式就有4種。鹼基對的排列順序就代表了遺傳信息。由此可見,DNA分子是能夠儲存大量的遺傳信息的。鹼基對的排列順序的千變萬化,構成了DNA分子的多樣性,而鹼基對的特定的排列順序,又構成了每一個DNA分子的特異性,這就從分子水平上說明了生物體具有多樣性和特異性的原因。DNA分子的複製DNA分子的結構不僅使DNA分子能夠儲存大量的遺傳信息,還使DNA分子能夠傳遞遺傳信息。遺傳信息的傳遞是通過DNA分子的複製來完成的。DNA分子的複製是指以親代DNA分子為模板合成子代DNA的過程。這一過程是在細胞有絲分裂的間期和減數第一次分裂的間期,隨著染色體的複製而完成的。
半保留複製
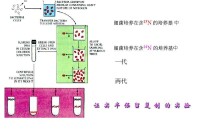
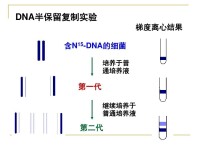
在上世紀中葉(1950s)JamesWatson和FrancisCrick提出了著名的DNA雙螺旋以及雙鏈間鹼基配對的模型,根據這個模型,他們進一步提出了DNA複製的半保留模型(semiconservativemodel),雖然這個模型比當時並存的全保留模型(conservative模型)看起來簡單易行的多,但始終缺乏有說服力的數據。最後在1957年,當時在Caltech作研究生的MatthewMeselson和作博士后的FranklinStahl設計並實現了這組著名的,證明了DNA複製半保留機理的實驗。試驗中,他們先將大腸桿菌細胞培養在用15NH4Cl作為唯一氮源的培養液里養很長時間(14代),使得細胞內所有的氮原子都以15N的形式存在(包括DNA分子里的氮原子)。這時再加入大大過量的14NH4Cl和各種14N的核苷酸分子,細菌從此開始攝入14N,因此所有既存的“老”DNA分子部分都應該是15N標記的,而新生的DNA則應該是未標記的。接下來他們讓細胞們繼續高高興興地生長,而自己則在在不同時間提取出DNA分子,利用CsCl密度梯度離心分離,最後得到每行從左到右由一個實驗代碼(這個數對我們來說沒啥用),一個小圖片,一個峰狀譜和一個被叫做generation的數字組成。最後這個數字實際上反映了從加入氮14開始,細胞進行了多少次分裂(也就是進行了多少次複製),左邊的黑色帶狀圖案反映了該樣品中DNA在離心管中的相應位置。大家可以想象一下這一列小圖片是一組疊置的離心管,每根管都是管口向左比齊(在密度梯度分離法中,密度越大的分子應該越接近管底,就上圖來說也就是越靠右)。這樣一來不難看出隨著細胞的分裂,上圖中DNA的密度有所減小,從而向左遷移。而當細胞分裂了一次的時候只有一個DNA帶,這就否定了所謂的全保留機理,因為根據全保留機理,DNA複製應該通過完全複製一個“老”DNA雙鏈分子而生成一個全新的DNA雙鏈分子,那麼當一次複製結束,應該一半DNA分子是全新(雙鏈都完全只含14N),另一半是“全老”(雙鏈都完全只含15N)。這樣一來應該在出現在離心管的不同位置,顯示出兩條黑帶。DNA帶的密度應當介於兩者之間,也就是相當於一根鏈是14N,另一根鏈是15N。而經歷過大約兩次複製后的DNA樣品(generation=1.9)在離心管中顯示出強度相同的兩條黑帶,一條的密度和generation=1時候的一樣,另一條則等同於完全是14N的DNA。這樣的結果跟半保留機理推測的結果完美吻合:就這樣,關於DNA複製機理的爭論終於被Meselson和Stahl完美解決,而基因學和基因組學也得以在此後的五十年取得一系列重大突破。
半保留複製
半保留複製
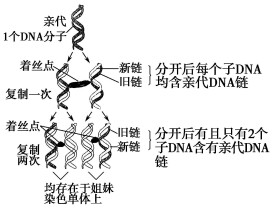
(2)染色體只有一半顯示放射性,而顯示放射性的每條染色體只有一條染色單體顯示放射性。
(一)第一代分裂后形成的每條染色體上的一個DNA分子兩條鏈一條有放射性一條沒有放射性,如果是半保留複製,以解旋以後的兩條鏈為模板(一條有放射性一條沒有放射性)在普通培養液中形成的兩條子鏈都沒有放射性,這樣複製后的每條染色體都有一條染色單體具有放射性(以有放射性的那條母鏈為模板形成的)一條染色單體沒有放射性(以沒有放射性的那條鏈為模板形成的),所以第一次分裂中期每條染色體上都有放射性,而每條染色體上的兩條染色單體一條有放射性一條沒有放射性。
(二)在第一代分裂的基礎上進行第二代分裂,由於第一代分裂中期的每條染色體上的染色單體一條有放射性一條沒有放射性,所以著絲點分裂后形成的染色體就只有一半具有放射性了,而有放射性的染色體其中的DNA只有一條鏈具有放射性,如果是半保留複製,以解旋后的兩條鏈為模板在普通培養液中形成兩條沒有放射性的子鏈,這樣有放射性的染色體複製后形成的兩條染色單體就只有一條染色單體顯示放射性。
基本信息
- 中文名
- 半保留複製
- 外文名
- semiconservative replication
- 提出時間
- 1927年
- 提出者
- (前蘇聯)尼古拉·科爾佐夫
- 應用學科
- 生物學
- 適用領域
- 真核生物