次生結構
次生結構
次生結構secondary structure裸子植物,大多數雙子葉植物和某些蕨類植物中,由根和莖的維管形成層和木栓形成層產生的結構。次生結構可增強植物體的支持、輸導和貯藏等功能。在植物演化過程中,對種子植物征服陸地起了重要作用。同時,研究次生生長的規律,在木材、橡膠、生漆、樹脂等利用上也具有重大的意義。
植物體產生次生結構使莖和根加粗的過程,稱為次生生長。
在次生生長中,由於維管形成層的活動,其衍生細胞向外分化形成次生韌皮部,向內形成次生木質部,共同組成次生維管組織。維管形成層的不斷活動,次生維管組織逐漸增多,從而使根和莖原來的外圍初生組織,如表皮、皮層受到擠壓。此時,在根中一般由中柱鞘、在莖中由表皮或皮層的細胞轉化為木栓形成層。木栓形成層向外產生木栓層,向內產生栓內層,共同組成周皮。周皮以外的表皮和皮層被擠毀,而由周皮代替了表皮起著保護作用。
通常在溫帶生長的裸子植物和木本雙子葉植物,由於維管形成層每年的周期性活動,在其莖和根內形成大量的次生結構。從外至內由周皮和次生維管組織組成,而其初生結構一般都被擠毀。單子葉植物一般缺少次生結構,但其中有些種類,如龍舌蘭屬植物等,它們可以由莖內維管束外圍的薄壁組織中發生形成層狀結構,產生次生維管組織。有些植物具有異常的次生生長,產生異常的次生結構。它們的形成層發生或活動不同於正常形成層,也有的形成層的位置正常,但所產生的次生木質部與次生韌皮部的分佈特殊;其形成層除正常地向外產生次生韌皮部外,在向內產生次生木質部的同時,也產生次生韌皮部,如馬錢屬。還有的在正常維管束外圍的薄壁組織中,連續產生多個同心圓式排列的形成層環,形成次生維管組織,如藜科植物。
初生木質部與初生韌皮部之間保留的一層具分裂能力的細胞發育為束中形成層,它構成了形成層的主要部分。此外,在與束中形成層相接的髓射線中的一層細胞,恢復分裂能力,發育為形成層的另一部分,因其位居維管束之間,故稱為束間形成層(interfascicular cambium)。束中形成層和束間形成層相互銜接后,形成完整的形成層環。
維管形成層細胞的組成有 紡錘狀原始細胞(fusiform initial)和 射線原始細胞(cray initial)兩種類型(圖5-37)。前者細胞長而扁,兩端尖斜,切向面比徑向面寬,它和莖的長軸平行排列;後者近乎等徑,分佈於紡錘狀原始細胞之間。在莖橫切面上,這種細胞都是呈平周的長方形,排成一圓環。
維管形成層分裂活動時,紡錘狀原始細胞進行平周分裂形成的新細胞向外逐漸分化為次生韌皮部,向內形成次生木質部,構成軸向的次生維管組織系統。紡錘狀原始細胞也可進行垂周分裂,增加自身細胞的數目以及衍生出新的射線原始細胞,從而使形成層環的周徑擴大。射線原始細胞平周分裂的結果,形成徑向排列的次生薄壁組織系統,即次生維管射線。其中,位於次生木質部中的部分稱為木射線;位於次生韌皮部的部分稱為韌皮射線,它們構成莖內橫向運輸系統。形成層活動過程中,往往形成數個次生木質部分子之後,才形成一個次生韌皮部分子,隨著次生木質部的較快增加,形成層的位置也逐漸向外推移。
形成層分裂活動所形成的次生木質部的量遠大於次生韌皮部的量。特別是木本植物的莖,絕大部分都是次生木質部,它擔負著輸導水分、無機鹽和支持功能。次生木質部的細胞組成與後生木質部的後期發育部分一樣,也包含導管、管胞、木纖維和木薄壁細胞。
⑴ 早材與晚材:維管形成層的活動易受外界環境條件的影響,在有明顯冷暖季節交替的溫帶,或有乾濕季節交替的熱帶,形成層的活動隨季節更替,表現出明顯的節奏性變化。形成層的活動有強有弱,形成的細胞有大有小,壁有厚有薄,顏色有深有淺,從而在次生木質部的形態結構上表現出明顯差異。在溫帶春季,氣候條件逐漸變暖,形成層活動也隨之增強,結果形成的次生木質部細胞多,其中導管和管胞的直徑大而壁薄,木材顏色較淺,木材質地較疏鬆,稱之為 早材(early wood)。在夏末秋初,氣候條件逐漸不適宜於樹木生長,形成層活動隨之減弱,形成的細胞數目減少,其中導管和管胞徑小而壁厚,木材顏色深,木材質地較緊密,稱之為 晚材(late wood)。
⑵ 年輪:年輪也叫 生長輪(growth ring)或 生長層(growth layer)。在一個生長季內,早材和晚材共同組成一輪顯著的同心環層,代表著一年中形成的次生木質部。在有顯著氣候變化的地區,植物的次生木質部在正常情況下,每年形成一輪,因此,習慣上稱為 年輪(annual ring)。但有不少植物在一年內的正常生長中,不只形成一個年輪,柑橘屬植物的莖,一年中可產生三個年輪,也就是三個年輪才能代表一年的生長,因此,又叫假年輪,即一個生長季內形成多年年輪。此外,氣候的異常,蟲害的發生,出現多次寒暖或葉落的交替,造成樹木內形成層活動盛衰的起伏,使樹木的生長時而受阻,時而復甦,都可能形成假年輪。沒有乾濕季節變化的熱帶地區,樹木的莖內一般不形成年輪。因此,年輪的數目通常可作為推斷樹木年齡的參考。
年輪還可以反映出樹木歷年生長情況,結合當地當時氣候條件和撫育管理措施的實際,進行比較和分析,可以從中總結出樹木快速生長的規律,便於指導林業生產,還可以從樹木年輪的變化中,了解到一地歷年及遠期氣候變化情況和規律。有的樹齡已達百年、千年之久以及地下深埋的具有年輪的樹木莖段化石,都是研究早期氣候、古氣候變遷的可貴依據。
⑶ 心材與邊材:多年生木本植物隨著年輪增多,莖幹不斷增粗。靠近樹皮部分的木材是近幾年形成的次生木質部,顏色較淺,導管有輸導功能,木薄壁細胞是活動的,稱為 邊材(sap wood)。而靠近中央部分的木材,是較早形成的次生木質部,顏色較深,其中,導管由於被其周圍薄壁細胞侵入,在導管腔中形成 侵填體(圖5-41),並積累單寧、樹脂、色素等物質,因而被堵塞,失去輸導能力,木薄壁細胞和木射線細胞也成為死細胞,此部分木材即為 心材(heart wood)。形成層每年形成新的次生木質部擴增於邊材的外側部分,而邊材的內側部分則逐漸轉變為心材,因此,邊材的數量較穩定,而心材的量逐年增加。
⑷ 木材解剖的三種切面:為了更好地理解莖的次生木質部,就必須從木材結構的三切面即 橫切面(sross section)、徑向切面(tangential section)和 切向切面(radial section)上對其進行比較觀察,從而建立立體模型,這樣,才能從立體的形象充分理解它的結構。橫切面是與莖的縱軸垂直所做的切面,可見到同心圓似的年輪,所見到的導管、管胞和木纖維等,都是它們的橫切面觀,可以看出它們細胞直徑的大小、壁厚、細胞形狀及細胞分佈狀況。所見的射線作輻射狀排列,這是射線的縱切面,可觀察到它們的長度和寬度;切向面也稱弦向切面,是垂直於莖半徑所做的縱切面,也就是離開莖中心所做的任何切面,可見到年輪呈U字形。在切向面上所見的導管、管胞、木纖維、木薄壁組織都是它們的縱切面,可觀察到它們的長度和寬度和細胞兩端的形狀。所見的射線是它們的橫切面,其輪廓為紡錘狀,顯示了射線的高度、寬度、細胞的列數和兩端的細胞形狀;徑向切面是通過莖直徑所做的縱切面,在徑向切面上,所見到的導管、管胞、木薄壁細胞、木纖維和射線都是它們的縱切面。細胞排列較整齊,尤其是射線細胞與莖縱軸垂直,長方形的細胞排列成多行,井然有序,象一段磚牆,顯示了射線的高度和長度。在三種切面中,射線形狀最為突出,以此可作為判別切面的指標。
次生韌皮部位於形成層外方,由篩管、伴胞、韌皮薄壁細胞和韌皮纖維組成,有的具有石細胞,杜仲(Eucommia)等植物的次生韌皮部中還有乳汁管分佈。形成層形成的次生韌皮部的數量遠較次生木質部的少。次生韌皮部維持輸導作用的時間較短,通常篩管只有1~2年的輸導能力。部分衰老的篩管由於篩板上形成胼胝體,堵塞篩孔,失去輸導作用,同時隨著次生生長的繼續進行,遠離形成層的先期產生的次生韌皮部,受到裡面增大的木質部的壓力也越來越大,篩管和一些薄壁細胞甚至被擠毀。當木栓形成層在次生韌皮部形成后,木栓形成層以外的韌皮部就完全死亡,成為乾死的組織而參與樹皮的形成。
木栓形成層的發生
部分草本雙子葉植物莖具有較弱的次生生長,其表皮細胞能進行細胞分裂,增加表皮周長,藉以適應莖幹的微緩增粗。木本植物莖幹的次生生長活躍,增粗顯著,表皮不能適應莖內的不斷增粗生長,以致最終死亡、脫落,而由木栓形成層產生的周皮代替了表皮的功能。
木栓形成層的活動
莖中木栓形成層的來源較為複雜,各種植物有所不同。多數植物的木栓形成層起源於與表皮鄰接的一層細胞(薄壁組織細胞或厚角組織細胞),但也有起源於表皮細胞(如柳樹、蘋果和夾竹桃等),還有的起源於初生韌皮部中的薄壁組織細胞(如葡萄、茶等)。
周皮形成時,枝條的外表同時形成一種通氣結構,稱為 皮孔(lenticel)。皮孔常發生於原先氣孔的位置,此處內方的木栓形成層不形成木栓細胞,而形成許多圓球形的、排列疏鬆的薄壁細胞,組成補充組織。由於補充組織的增加,向外突起,將表皮脹破,形成裂口,即為皮孔。皮孔的形成改善了老莖的通氣狀況。
有些植物的木栓形成層的壽命較長,最初形成的木栓形成層可以保持多年甚至終生不失其效能(如栓皮櫟)。但多數植物木栓形成層的壽命較短,在短時期內木栓形成層本身也轉變成木栓組織了,在此情況下,在它內方發生出新的木栓形成層,再形成新周皮。隨著莖的增粗,木栓形成層發生的部位逐漸向內推移,甚至可達次生韌皮部。新周皮形成后,其外方所有的活組織由於得不到養料和水分的供應以及被擠壓而死亡,這些失活的組織,包括多次形成的周皮以及周皮以外的死亡組織組成了樹皮(脫落皮層或樹皮),但也有將形成層以外的所有組織統稱為樹皮的,這是樹皮的廣義概念。另外樹皮色澤、形狀以及皮孔和芽的形態特徵常依植物不同而有差別,可作為鑒別冬季落葉樹種的依據。
大多數雙子葉植物和裸子植物莖完成初生生長后,由於次生分生組織的活動,使莖不斷增粗,這種增粗生長稱為次生生長,也稱加粗生長。次生生長所形成的次生組織組成了次生結構。多年生木本植物,不斷地增粗和增高,必然需要更多的水分和營養,同時,也需要更大的機械支持力,因此必須相應地增粗。次生結構的形成和不斷發展,才能滿足多年生木本植物在生長和發育上的這些要求,這也正是植物長期生活過程中產生的適應性。
單子葉植物莖的次生結構
大多數單子葉植物沒有次生生長,因而也就沒有次生結構。莖的增粗是由於細胞的長大或初生加厚分生組織平周分裂的結果。但少數熱帶或亞熱帶的單子葉植物莖,除一般初生結構外,有次生生長和次生結構出現,如龍血樹、朱蕉、絲蘭、蘆薈等的莖中,它們的維管形成層的發生和活動,不同於雙子葉植物,一般是在初生維管組織外方產生形成層,形成新的維管組織(次生維管束),因植物不同而有各種排列方式。現以龍血樹(Dracaena draco)為例,加以說明。
龍血樹莖內,在維管束外方的薄壁組織細胞能轉化成形成層,它們進行切向分裂,向外產生少量的薄壁組織細胞,向內產生一圈基本組織,在這一圈組織中,有一部分細胞直徑較小,細胞較長,並且成束出現,將來能分化形成次生維管束。這些次生維管束也是散生的,比初生的更密,在結構上不同於初生維管束,因為所含韌皮部的量較少,木質部由管胞組成,並包於韌皮部的外周,形成周木維管束。而初生維管束為外韌維管束,木質部由導管組成。
裸子植物莖的次生結構
?
裸子植物都是木本植物,莖的結構基本和雙子葉植物木本莖大致相同,二者都是由表皮、皮層和維管柱等部分組成,長期存在著形成層,產生次生結構,使莖逐年加粗,並有顯著的年輪,不同之處是維管組織的組成成分上存在差異。
⒈多數裸子植物莖的次生木質部是由管胞和射線組成,無導管和典型的木纖維,管胞兼有輸送水分和支持的雙重作用。在橫切面上,結構顯得均勻整齊。和雙子葉植物莖的次生木質部相同,裸子植物的次生木質部中也存在著早材與晚材,邊材與心材的分化。
⒉裸子植物次生韌皮部的結構較簡單,由篩胞、韌皮薄壁組織和射線組成,沒有篩管、伴胞和韌皮纖維。有些松柏類植物莖的次生韌皮部中,也可能產生韌皮纖維和石細胞,有些種類可以產生樹脂道,如松。
禾本科植物莖節次生結構
禾本科植物的莖和葉鞘相連形成了節部。在外形上,節部比較粗大,易於識別。內部結構上,由於上端節間維管束以及從葉鞘伸入的維管束(葉跡)在此交織匯合,出現了比較複雜的結構。將小麥莖,由上至下,從上部節間經節部,再到下部節間作連續切片觀察,可以看到這種維管系統匯合排列的變化過程(圖5-49)。
小麥莖的節間中空,在節部成為實心。葉鞘在較高水平上向一邊開放,到了靠近節部成為封閉的。內部結構上,莖節上面的維管束成橫向和斜向的分佈,而在節內以及節的下面,就重新組合排列到較外面的周圍。在葉鞘和莖連接處的下面,較小的葉跡伸到莖軸的外圍,較大的葉跡則變成了莖軸的中間維管柱的一部分。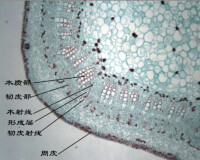
次生結構

基本信息
- 中文名
- 次生結構
- 外文名
- secondary structure
- 拼音
- cì shēng jié gòu
- 類別
- 裸子植物
- 作用
- 科學研究