DNA甲基化
表觀遺傳調控機制之一
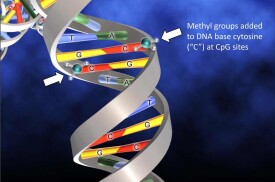
DNA甲基化(DNA methylation)為DNA化學修飾的一種形式,能夠在不改變DNA序列的前提下,改變遺傳表現。所謂DNA甲基化是指在DNA甲基化轉移酶的作用下,在基因組CpG二核苷酸的胞嘧啶5'碳位共價鍵結合一個甲基基團。大量研究表明,DNA甲基化能引起染色質結構、DNA構象、DNA穩定性及DNA與蛋白質相互作用方式的改變,從而控制基因表達。
DNA甲基化是最早被發現、也是目前研究最深入的表觀遺傳調控機制之一。廣義上的DNA甲基化是指DNA序列上特定的鹼基在DNA甲基轉移酶(DNA methyltransferase,DNMT)的催化作用下,以s一腺苷甲硫氨酸(S—adenosyl methionine,SAM)作為甲基供體,通過共價鍵結合的方式獲得一個甲基基團的化學修飾過程。這種DNA甲基化修飾可以發生在胞嘧啶的C一5位、腺嘌呤的N一6位及鳥嘌呤的N一7位等位點。一般研究中所涉及的DNA甲基化主要是指發生在CpG二核苷酸中胞嘧啶上第5位碳原子的甲基化過程,其產物稱為5—甲基胞嘧啶(5一mC),是植物、動物等真核生物DNA甲基化的主要形式,也是目前發現的哺乳動物DNA甲基化的唯一形式。DNA甲基化作為一種對穩定的修飾狀態,在DNA甲基轉移酶的作用下,可隨DNA的複製過程遺傳給新生的子代DNA,是一種重要的表觀遺傳機制。
基因組中DNA的甲基化模式是通過DNA甲基轉移酶實現的。DNA甲基化酶分為2類,即維持DNA甲基化轉移酶(Dnmtl或維持甲基化酶)和從頭甲基化酶。根據序列的同源性和功能,真核生物DNA甲基化轉移酶又分為4類:Dnmtl/METl、Dnmt2、CMTs和Dn-mt3。DnmtliiMETl類酶參與CG序列甲基化的維持。CMTs類酶僅發現在植物中,主要特徵是它的催化區T和Ⅳ包埋染色體的主區,並且特異性地維持CG序列的甲基化。Dnmt:3類酶在小鼠、人類和斑馬魚中得到鑒定.Dnmt3a和Dnmt3b在未分化的胚胎幹細胞中高度表達,但在體細胞中表達水平很低。它們的主要作用是從頭甲基化,但對維持甲基化也起到一定的作用,並且負責重複序列的甲基化。
DNA甲基化反應分為2種類型。一種是2條鏈均未甲基化的DNA被甲基化,稱為從頭甲基化(denovo methylation);另一種是雙鏈DNA的其中一條鏈已存在甲基化,另一條未甲基化的鏈被甲基化,這種類型稱為保留甲基化(maintenance methylation)。
由於Dnmtl和Dnmt3基因家族沒有針對CpG二核苷酸序列的特異性,人們因此提出了DNA甲基化轉移酶發現靶位點的機制。首先,甲基化轉移酶並不是同等地接近所有染色體區域。具有染色體重構和DNA螺旋酶活性的蛋白質能調節哺乳動物細胞內DNA甲基化,如SNF2家族2個成員ATRX和Lsh;其次,附件因子(蛋白質、RNA等)能召集DNA甲基化轉移酶到特定基因組序列或染色體結構中,如pRB蛋白等能夠與Dnmtl作用,在S期晚期將它召集到高度甲基化的異染色質區。
DNA甲基化(methylation)是真核細胞正常而普遍的修飾方式,也是哺乳動物基因表達凋控的主要表觀遺傳學形式。DNA甲基化后核苷酸順序及其組成雖未發生改變,但基因表達受影響。儘管甲基化修飾有多種方式,被修飾位點的鹼基可以是腺嘌呤的N-6位、胞嘧啶的N-4位、鳥嘌呤的N-7位和胞嘧啶的C-5位,它們分別由不同的DNA甲基化酶催化,但大多發生在基因啟動子區CpG島上。DNA甲基化時,胞嘧啶從DNA雙螺旋上突出,進入能與酶結合的裂隙中,在胞嘧啶甲基轉移酶催化下,把活性的甲基從S一腺苷甲硫氨酸轉移至胞嘧啶5位上,形成5一甲基胞嘧啶(5一MC)。基因啟動子區的甲基化可導致轉錄沉寂。
哺乳類動物一生中DNA甲基化水平有著顯著變化:在受精卵最初幾次卵裂中,去甲基化酶清除DNA分子上幾乎所有從親代遺傳下來的甲基化標記;在胚胎植入子宮時,一種新的甲基化遍布整個基因組,構建性甲基化酶使DJA重新建立一個新的甲基化模式,一旦細胞內新的甲基化模式建成,將通過維持甲基化酶以“甲基化維持”的形式將新的DNA甲基化模式傳遞給所有子細胞DNA分子上。這就解釋了基因印記不是一種突變,也不是一種永久的變化。印記是可逆的,它只持續於個體的一生中,在下一代個體的配子形成時,舊的基因印記消除並又發生新的基因印記。由此可見,遺傳印記的分子機制之一可能就是DNA甲基化。在遺傳印記與腫瘤的研究中也發現腫瘤抑制基因P16,甲基化使之失活,去甲基化可使之恢復該基因本來的特性。而異常甲基化可能是腫瘤發生的重要原因之一。說明DNA甲基化的修飾有著廣泛的作用。
甲基化的DNA可以發生去甲基化。DNA的去甲基化由基因內部的片段及與其結合的因子所調控。有兩種假說可以解釋DNA去甲基化的分子機制。一種假說與DNA半保留複製聯繫在一起,為被動去甲基化。如果甲基化的DNA經半保留複製后不被甲基化,其DNA則處於半甲基化狀態,半甲基化的DNA如再次發生DNA半保留複製,而DNA甲基化活性仍被抑制,則有50%細胞處於半甲基化狀態。第二種假說與半保留複製無關,為主動過程。DNA去甲基化由DNA去甲基化酶催化。DNA去甲基化是在DNA糖苷酶的作用下脫掉甲基化鹼基的反應,等同於被損傷的DNA在糖苷酶及無鹼基核酸酶酶切偶聯催化下的修復反應。5一甲基胞嘧啶糖基化酶是體內侯選去甲基化酶。此外,甲基化CpG結合蛋白如MBD2等也具有去甲基化酶的活性。
細胞發育過程中,各種表觀遺傳學現象之間不是孤立存在而是密切聯繫的。DNA甲基化同組蛋白甲基化共同調控基因表達的現象最早在鏈孢霉(Neurospora crassa)中得到證實,進一步的生化研究結果表明,DNA甲基化是受組蛋白甲基化調節的。對哺乳動物的研究發現,DNA甲基化是建立和維持其他表觀遺傳學現象的基礎,比如DNA甲基化位點可以募集諸如組蛋白去乙醯化酶等具有抑制功能的複合物,同時除掉該位點附近的組蛋白乙醯化標記。也有研究認為,DIA甲基化是受組蛋白修飾調控的,有報道稱組蛋白修飾H3K9me能夠促進DNA甲基化的進程。