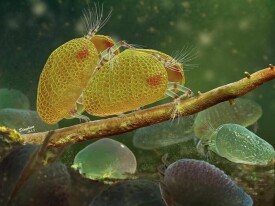
介形蟲
軀幹短、外表不分節的甲殼動物
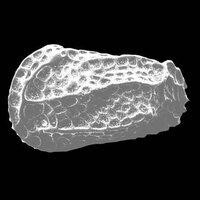
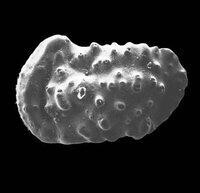
介形蟲由大小相等或不等的兩殼瓣組成,兩殼均勻向外凸出,殼面上光滑或有花紋、刺狀突起和槽等。殼體俯視呈卵形、紡錘形、馬蹄形等,殼長0.5~4 mm,最大可達30 mm。介形蟲殼主要由幾丁質組成,鈣化或磷化限於殼外層,並由外向內進行。介形蟲以其玻纖結構、殼小、邊緣具疊復現象與其他雙殼化石相區別。介形蟲可以生活於各種水的環境中,大多數是海相的,也有廣鹽性、微鹹水或超鹹水的湖或某些淡水中。時代為寒武紀—現代。

介形蟲
介形蟲綱( )甲殼,布廣。似軟體動物的兩瓣外殼。軀幹極短,外表不分節,這與多數甲殼動物不同。幾乎整個身體包在外殼內。據估計,現生種數量大約在1萬到1.5萬之間,包括海生、淡水生和陸生種類。已知的滅絕的和現生的介形蟲共有65,000種,從早奧陶世到近代已發現許多化石。由於其數量多,地理分佈廣,因此在地層中留下豐富的化石,可作為地層的指示化石。有的屬通常用來指示地下石油的勘探,與海洋地層中有孔蟲的情形有些類似。現生介形蟲的成蟲約0.3-5毫米長,太平洋最大的大介蟲(Gigantocypris agassizi)長23毫米(約1寸)。在海洋中,多數種生活在近海底處。在陸地上的湖泊、沼澤、池塘、河流、甚至土壤間隙水(例如水井)中都有分佈。有的吃微生物和有機碎屑;有的吃小型無脊椎動物;少數寄生。多數喜歡在淺水中,但有的采自2,000公尺(約6,600尺)深處。
介形蟲
被包在兩瓣殼內。殼形多樣。軀體兩側對稱,不分節,被包在兩個殼瓣之間。殼面光滑或具各種瘤、槽、刺、脊以及其它紋飾,但不具生長線(這是與軟體動物外殼的區別)。器官發育較完善,身體分頭胸部和腹部。具附肢,末端具尾叉。兩殼瓣可不相等,殼質分兩層外層--鈣質層;內層--幾丁質,邊緣可鈣化。有的種具雌雄雙形現象。生活領域廣泛,能適應淡水、海水等多種生活環境。自早奧陶世出現,一直到現代還相當繁盛。(也有觀點認為出現於寒武紀。爭論的焦點在於高肌介類是否屬於介形蟲。)
分類
節肢動物門(Arthropoda)甲殼超綱(Crustacea)
介形蟲綱(Ostracoda)
高肌介目(Bradoriida,目前認為不是介形蟲)
豆石介目(Leperditicopida,目前認為不是介形蟲)
古足介目(Palaeocopida)
速足介目(Podocopida)
麗足介目(Myodocopida)
介殼化石的分類依據
1、介殼形狀
2、殼面裝飾
3、肌肉印痕
4、兩殼的疊覆關係
5、鉸合構造(很少用)
6、內邊緣構造
7、眼點
介形蟲的近親之一,高肌介類從寒武紀早期出現,化石分佈於亞、歐、澳及北美地區。但目前的主流觀點認為高肌介不屬於介形蟲。目前已知的最早出現的介形蟲,發現於距今4.85億年前的早奧陶世地層中的古足介類。奧陶紀是古足介目全盛時期。三疊紀時速足介目開始占統治地位。侏羅紀時,浪花介超科、金星介超科、小浪花介科等大量繁盛。同時,達爾文介超科、土菱介超科等仍繼續發展。白堊紀開始,介形蟲動物群再次出現大發展時期。更新世至現代介形蟲屬的數量巨大。
生物地層意義:中新生代海陸相地層中介形蟲可分為很多組合,具有重要的地層學意義。
科學家首次發現5.4億年前生活在地球上的介形蟲的巨型精子化石,這是地球上已知最古老生物的生殖細胞。
生物的軟體組織經過數百萬年的時間通常不易保存。但這次研究人員很幸運,他們發現的化石以貝殼的形狀,保存在一塊琥珀中。樹脂本身也成為化石,但研究人員成功將琥珀熔化。研究發現,這塊琥珀中包含39個幾乎完整的介形蟲。
據科學家在《皇家學會會刊》上發表的一篇文章,這些介形蟲中包括雄性、雌性和幼蟲。軟體組織和精子都被保存下來,年代可以追溯到1億年前。
古生物學家研究對比了此次發現的介形蟲和目前生存的介形蟲,並藉助X光獲得高解析度的三維圖像。科研人員最終發現了雌性的儲精囊和生殖細胞本身。
主要構造

介形蟲介殼外視
尾叉。眼位於頭部背側,通常為單個的眼,多數介形蟲不具眼,有的屬種幼年期具眼,到成年期消失。介形蟲身體某些部位生有感覺毛,通過殼壁的毛細血管伸出殼外司感覺作用。消化系統位於頭部腹側的口、食道、胃、腸及位於身體後端的肛門組成。無腮,通過薄的體壁自然擴散進行呼吸。除了個體較大,浮遊類型的目之外,均缺少血管和心臟。肌肉系統複雜。

介形蟲
型起源、但外肢退化,主要用於爬行、挖掘、進食以及交配。
雌雄異體。有的種類無雄性,以單性生殖方式產生後代,有些屬種行兩性生殖,也有兩性生殖和單性生殖都可以進行。雌性生殖器官由位於身體後部的卵巢及輸卵管組成。在特定季節生殖。卵呈圓形或卵圓形,耐乾旱及寒冷性很強,利於適應低溫、乾旱環境,並可較長距離擴散,在適宜的溫度下孵出幼蟲。它的增長是不連續的由卵孵化出幼蟲稱第一齡期,當身體生長對殼體來說已經過大時,堅硬的雙殼脫落,隨之身體迅速增長和發育,同時形成新的殼並硬化。由第一期至成年,通常經過8個這樣的齡期,蛻殼8次,每蛻殼一次,個體增大一些,各器官、附肢不斷增加、完善,最後達到成年。期發育過程中,殼形、殼飾及各種構造特徵也發生一定規律性的明顯變化。
根據運動前進方向分左、右兩瓣

介形蟲?
上表皮、外表皮、內表皮和表皮細胞四層。上表皮極薄,不能保存為化石;外表皮為鈣質。常保存為化石;內表皮也為鈣質;表皮即由節肢動物的表皮細胞組成。內殼層由幾丁質組成,甚薄,僅邊緣部分鈣化成較厚的鈣化擘,起著增進殼的自由邊緣閉合的作用。幼年期和某些淡水介形蟲及浮遊的麗足介目的大多數種屬的殼質鈣化弱,不一保存為化石。
介形蟲?
具各種不同紋飾。位於前背部的圓形小結節或小坑稱眼痕。具雙形現象,即同一種的雌雄不同殼體的形態特徵往往存在顯著差別。
殼內側構造:鉸合構造--殼內側、背部邊緣、突起的齒、脊和凹入的齒窩、槽組成
肌肉印痕--閉殼肌痕、大額肌痕
邊緣構造--內緣板、結合帶、結合線、毛細管帶
這些都具有重要的鑒定意義
1、研究介形蟲種屬的分類、組合、丰度及分異度、殼體的大小、形態、結構、殼飾及厚度;
2、分析水化學條件: 鹽度、離子組成、溫度、深度、食物的供給等;
3、水體的溫度、鹽度、酸鹼度影響介形蟲生態特徵;
4、介形蟲的古生態學,可以提供重要的環境變化信息,掌握鹽度和離子組成對介形蟲種屬變化的影響;
5、了解介形蟲生命歷史和生態消長的過程。
要想測量海洋的深淺,除了使用各種測深儀實際測量外,還可從介形蟲得到大海深淺的數據。介形蟲有自己固定的棲息地,從不到處漂泊,例如在深海生活的種類決不到淺海處棲居;在淺海處生活的種類也決不到深海去邀游。地質學家根據介形蟲的這一習性,就能估算出大海的深淺。例如,在我國南黃海西北部地區海底泥沙中介形蟲的分佈,南部以中華麗花介為主,北部以穆賽介為主,東部以克利特介為主。這三種介形蟲分別生活在0-20m、20-50m和大於50m水深的海區。因此,根據這些介形蟲種類的分佈情況,就能繪製出一幅簡單的海底地形圖。
對於現代的海洋測量來說,介形蟲給出的深淺數據當然是太粗略了,根本無法與現代儀器的精密測量相提並論。但是,介形蟲具有測量千百萬年前海水深淺的本領,卻是任何現代精密測深儀都望塵莫及的。現代測深儀無論多麼先進,只能測現代海洋的深淺,對於遙遠地質年代的海洋的深淺則無能為力。在漫長的歷史進程中,海洋早已發生了巨大的變化。面對這個面目全非的海洋,介形蟲卻能大顯身手。比如,地質學家從地中海幾千萬年前形成的沉積物中,發現了一種叫深海角介的只能在大洋里生活的介形蟲,而在年代更新的沉積物中,卻再也見不到它的蹤跡。由此得知,古地中海曾經是一個大海,與大西洋相通,水深可能達到幾千米,以後它又與大洋失去聯繫,封閉成如今名副其實的被陸地包圍著的地中海。在這一點上,介形蟲所提供的寶貴數據是無與倫比的。

介形蟲
據悉,這一精子是在一隻已經石化了的雌性遠古物種介形蟲的性器官內發現的,科學家稱這隻介形蟲在死前剛剛交配過。相對於這種動物的體型而言,該精子顯得非常大。
精子藏身的化石由三名研究人員於數年前發掘。其中一名研究員阿徹教授(Michael Archer)表示,99.9%微生物的精子與體積相比都非常小,而且這隻介形蟲的軟組織能保存得如此完好,令我們感到異常驚訝。
“當撿起這些堅硬的、乾燥的石頭時,我們通常會用鎚子敲它。我們完全沒有想過在這些石頭裡會發現曾存在於軟組織內的精子。”阿徹教授說。
阿徹教授亦指出,目前尚不清楚這隻介形蟲的細胞是如何變成化石的。但蝙蝠分泌出來的液體在保存精子的過程中可能發揮了重大作用。而蝙蝠液體大量存在於發現化石的洞穴地面。
1. 簡介
介形蟲是一類小型甲殼動物,屬於節肢動物門、甲殼動物亞門、介形蟲綱。它們的整個身體包被在一對外殼中。外殼的長度一般介於0.3–5 mm之間,主要成分通常是方解石和幾丁質。身體由頭、胸、腹部構成,但體表不可見分節。通常具8對附肢,除第一觸角外,均為雙枝型起源。現生介形蟲廣泛分佈於各種水域。目前發現的最早的介形蟲化石屬於4.85億年前的奧陶紀。
註:1)由拉丁文Ostracoda派生出介形蟲的兩個英文詞:ostracod是除法國以外的歐洲、以及澳洲的寫法,ostracode是法國及北美的寫法。拉丁文Ostracoda的首字母須大寫,是分類學的用法,表示介形蟲綱。
2)過去,人們為甲殼動物設甲殼綱,為介形蟲設亞綱。近年始變。
3)介形蟲的附肢自前而後有:第一觸角、第二觸角、大顎、小顎、第一胸肢、第二胸肢、第三胸肢、尾肢。由於體表不可見分節,因此所謂的第一胸肢屬胸部還是頭部尚有爭議。
4)介形蟲的第一觸角是單枝型的,其餘觸角是雙枝型的,儘管雙枝型的部分特徵有時退化至不可見。這與其它甲殼動物一致。雙枝型附肢的基本構造包括原肢(protopod)以及由原肢分出的外肢(exopod)和內肢(endopod)。此外,原肢上有時還著生有內葉(endite)和上肢(epipod)等結構。
5)尾肢(uropod)曾被稱為furca,並被認為起源於身體的尾節(telson)。但現在認為它起源於腹部一對附肢的振動片(vibratory plate)(Meisch, 2007)。它也是多數介形蟲唯一的腹部附肢。在某些浪花介超科的雄性中,尾肢之前還有一對稱為brush organ的附肢。
6)上面所說的8對附肢不包括由附肢演化而來的陰莖等結構。
7)目前公認的最早的介形蟲是出現於奧陶紀早奧陶世的古足介類(Palaeocopida)。出現於寒武紀的高肌介類(Bradoriida)不被視作介形蟲。詳見下文。
2. 分類系統
如圖1所示,介形蟲綱下分四個目:古足介目Order Palaeocopida Henningsmoen 1953,麗足介目Order Myodocopida Sars 1866,平足介目Order Platycopida Sars 1866,以及速足介目Order Podocopida Sars 1866。高肌介目Bradoriida和豆石介目Leperditicopida。是否應划入介形蟲綱,還存在較大的爭議,原因就在於,這兩類動物都已經滅絕,留下的化石大多是外殼,很難找到保存良好的軟體,從而根據附肢的解剖結構來判斷其分類地位。從目前發現的唯一的高肌介軟體化石、來自雲南澄江寒武系的Kunmingella douvillei(Hou et al., 1996)來看,其附肢結構不同於普通介形類(Williams et al., 2008)。因此,目前的主流學派把高肌介排除在介形蟲綱之外。至於豆石介,筆者不清楚是否有軟體發現,但介形蟲學界似乎也傾向於不將它納入介形蟲。
上面的這個爭議也引發了另外一個問題,即最早的介形蟲到底是出現於寒武紀還是奧陶紀。因為高肌介出現在寒武紀,而目前公認的屬於介形蟲的四個目中,最早出現的是古足介目——發現於距今4.85億年的早奧陶世地層中(Williams et al., 2008)。如果高肌介屬於介形蟲,那麼就應該說,最早的介形蟲出現於寒武紀;如果高肌介不是介形蟲,那麼介形蟲則應起源於奧陶紀。由於一般認為高肌介不屬於介形蟲,所以,一般也認為,介形蟲起源於奧陶紀、而不是寒武紀。
但是,值得再次強調的是,高肌介和豆石介的分類地位是懸而未決的問題。以高肌介目前僅有的附肢化石證據,即Kunmingella douvillei化石,尚不足以將所有的高肌介排除在介形類以外。隨著新化石的發現,將來很有可能在地層中發現屬於介形類的其它的高肌介軟體化石。分子生物學證據也暗示,介形類的起源很可能比目前發現的4.85億年要早得多(Regier et al., 2005)。因此,目前所發現的4.85億年前的古足介,很可能不是最早的介形蟲;更早的介形蟲可能另有其“蟲”。
根據Ostracon社區里介形蟲學家的推測,目前地球上生活著大約10000到15000種現生介形蟲,其中大約三分之二在海洋里,三分之一在各種陸地水域中。在現生介形蟲的三個目中,速足介最為繁盛。目前我們在各種陸地水域中發現的介形蟲,幾乎無一例外地屬於速足介。速足介目又分為三個超科(圖1):達爾文介超科Superfamily Darwinuloidea Brady & Norman 1889,金星介超科Superfamily Cypridoidea Baird 1845和浪花介超科Superfamily Cytheroidea Baird 1850。這三個超科中又以金星介種類最多——至少在陸地水域中是這樣。
在金星介超科中,玻璃介科Family Candonidae Kaufmann 1900和金星介科Family Cyprididae Baird 1845是大科。就非海洋種類來說,玻璃介科已知的種類有大約550種,金星介科有大約1000種,占所有三個超科非海洋介形蟲種類的75%(Martens et al., 2008)。

1

圖2
3. 軟體形態結構
介形蟲可以分為外殼和軟體兩部分。外殼主要是方解石,可在地層中保留成為化石,為古生物學家、地層學家、古氣候學家所感興趣;軟體分為身體和附肢,以及由附肢所演化而形成的陰莖等器官,是現代生物學家研究的對象。現生介形蟲分類的主要依據是其成蟲附肢的形態結構,而化石介形蟲的分類,則幾乎只能依據外殼。依據外殼進行的屬種鑒定有一定的不確定性,尤其是對於玻璃介科、土星介屬等類群。下面以金星介科異星介屬、土星介科土星介屬、以及玻璃介科豆形玻璃介屬為例,簡要介紹介形蟲的軟體結構。所有標本均為筆者采自國內的成蟲,所有照片均為筆者拍攝。下文描述的結構模式,除加以說明外,僅適用於金星介科。
1)第一觸角,antennule (A1),見圖3
分為8節(圖3)。第一和第二節融合,因此有人又把這兩節合稱第一節,而把第一觸角分為7節。與其它甲殼動物一樣,介形蟲的第一觸角在起源上可能不同於其它附肢,因為它是單枝的(uniramous),沒有內肢(endopod)和外肢(exopod)之分;而其餘附肢是雙肢(biramous)的,有內肢和外肢之分。
大多數金星介超科種類在每節的剛毛數目和位置上都是一致的。但其中某些豆形玻璃介的第一觸角剛毛式則有所不同:它在第一節上多出一根背毛,第三節、第四節、第五節、第六節各少一根腹毛,第七節也少一根毛。另外,至於每根毛的長短、粗細,以及其上是否著生小毛,也因屬種而異。
第一觸角的剛毛式在金星介科的不同介形蟲之間被認為是比較保守的,因此很少有人用它來對介形蟲進行分類。但實際上,某些毛的長度或可用作介形蟲分類的依據。
第一觸角主要用於運動(游泳、鑽掘等)和感知。
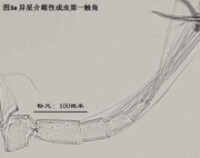
圖3a
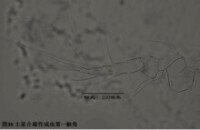
圖3b
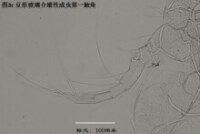
圖3c
2)第二觸角,antenna (A2),見圖4
與其它甲殼動物一樣,介形蟲的第二觸角是雙枝的,有內、外肢之分,它們是著生在原肢(protopod)上的兩個分枝,內肢靠內,外肢靠外。其中外肢退化為一長兩短三根剛毛,而內肢則粗壯,構成附肢的主幹。
第二觸角共有6節,其中原肢有2節、相互融合,內肢有3-4節。許多種類的第二觸角形態存在顯著性別分化。第二觸角上著生的大多數剛毛和爪子(爪子可認為由剛毛演化而來)比較保守,但第3節末端內側著生的6根游泳剛毛的長度,第4a節末端內側著生的4根t系列剛毛,以及第4b和5節末端著生的z系列的剛毛(某些種類雄性的z1和z2剛毛變為爪子)和G系列的爪子(某些種類雄性的G3細小,呈剛毛狀)的形態,是較為重要的分類學特徵。
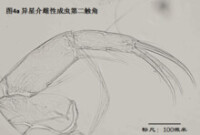
圖4a
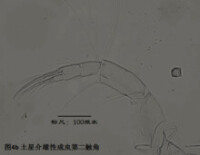
圖4b
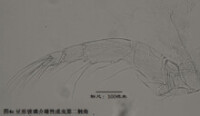
圖4c
3)大顎,mandible (Md),見圖5
大顎由coxa(masticatory process),振動片(vibratory plate)以及palp組成。其中coxa相當於原肢,vibratory plate由外肢演化而來(Karanovic, 2012),palp則相當於內肢。通常認為,內肢的剛毛式是重要的分類學依據,尤其是其S系列的剛毛(著生於內肢第一節),以及α,β,γ(分別著生在內肢的第一、第二和第三節)剛毛的形態。但實際上大顎的剛毛很多,常重疊在一起,觀察起來很費勁。
大顎用於攝食、咀嚼,有的種的大顎也用於運動。

圖5a
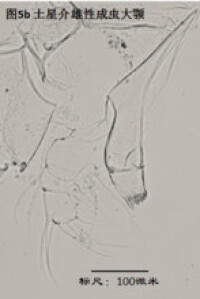
圖5b
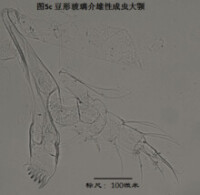
圖5c
4)小顎,maxillule (Mxl),見圖6
基本構成與大顎相似,由endite(可能相當於大顎coxa上的牙齒),稱為palp、分2節或不分節的內肢,以及由外肢演化而來的振動片(Horne, 2005)組成。
小顎用於攝食、咀嚼。其振動片較為發育,用於划動水流、從而使機體獲得足夠的氧氣。
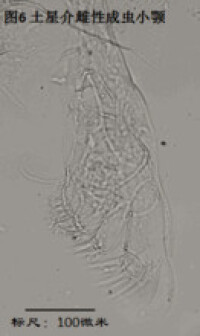
圖6
基本構成與前兩對附肢相似。由masticatory process(可能相當於endite),分節或不分節的palp,以及振動片組成。endite位於前方,palp和振動片位於後方。
雌性第一胸肢沒什麼可說的。用於攝食(浪花介超科的用於行走)。
雄性第一胸肢的palp,在許多金星介超科中特化為執握器(clasping organ,或prehensile palp),常較為粗壯,用於在交配中抓住雌性。左右執握器常不對稱。由於執握器是與生殖相關的器官,因此不同種的執握器常差異明顯。有經驗的介形蟲學家有時僅僅根據執握器就能鑒定介形蟲的種。

圖7
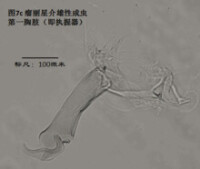
圖7-2
6)第二胸肢,the second thoracopod (T2或L6),見圖8
在所有速足目介形蟲中,這對附肢都是用來走路的,稱為步足(the walking leg)。共有6節。原肢實際上是2節融合在一起,每節的前側的末端分別著生一根毛,分別稱為d1和d2。筆者不清楚d2是否是外肢。內肢共有4節,但有時第二、第三節內肢(即第二胸肢的第四、第五節)相互融合。這種情況與第二觸角有些類似;但融合併非由於性別的緣故,而是在某些種的雌雄都如此,例如隆起土星介(Ilyocypris gibba)。內肢的每一節,不論它與相鄰的節融合與否,在其前側的末端都著生有一根有名字的毛:e,f,g和h1。倒數第二節,除了g外,還有一根沒有名字的毛。在最末一節,除h1外,還著生有一根粗壯的爪子h2和后側的毛h3。最末一節通常為一爪兩毛,偶爾是兩爪一毛。
步足的剛毛數目、位置和長短是重要的分類學依據。
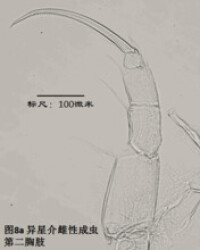
圖8a
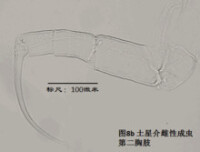
圖8b
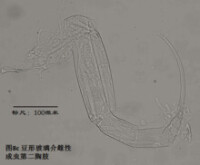
圖8c
7)第三胸肢,the third thoracopod (T3或L7),見圖9
其結構與第二胸肢明顯是同源的,但原肢末端多了一根毛,dp。金星介超科的第三胸肢向後、向上蜷曲,特化為一對清潔足(the cleaning leg),用於清潔介殼內部。兩節原肢的融合線常不可見。與第二胸肢類似,第三胸肢的時第二、第三節內肢(即第四、第五節)常完全融合以至於融合線不可見。在金星介科中,第四五六節完全融合,並且g和h2剛毛特化,h1很短,末端形成一個“pincer organ”(圖9a)。
注意:由於步足和清潔足的融合很完全,通常無痕迹,並且在同一種的兩性間是一致的,因此習慣上把融合的節合稱一節;這與第二觸角4a和4b即使融合仍分開標註的習慣不同。(由於第二觸角僅在某些種的雄性中有分界線,雌性中都是融合的,因此如果不分開標註,就會造成雌雄個體對比描述上的混亂。)
清潔足的剛毛數目、位置和長短也是重要的分類學依據。
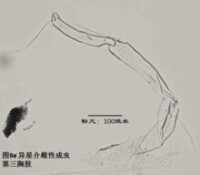
圖9a
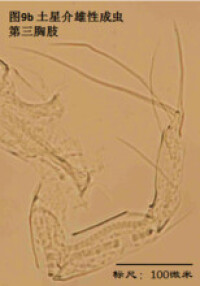
圖9b
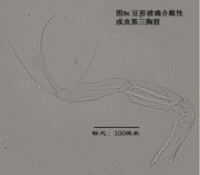
圖9c
8)尾肢,uropod (Ur),見圖10
由一根不分節的主幹(uropodal ramus,複數rami),兩個爪Ga和Gp,以及兩根毛Sa和Sp組成。a意為anterior,前方,顧名思義,Ga和Sa是位於前面的;p意為posterior,後方,也就是說Gp和Sp是位於后側的。
ps: 介形蟲的尾肢通常向下懸著,並斜指前下方。
尾肢主要用於運動(游泳或行走)。
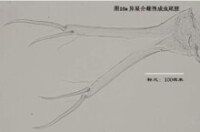
圖10a
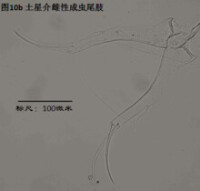
圖10b

圖10c
9)生殖器官。
雌、雄的外生殖器都成對出現,並且都位於第三胸肢和尾肢之間。
雄性陰莖,penis,更嚴格的提法是hemipenis (Hp),指一對陰莖中的一個,見圖11
與我們通常所認識的“陰莖”的概念有所不同,介形蟲的陰莖不光包括用於插入雌性外生殖器的部分,還包括很多其它的結構。但這些結構的作用知之尚少(Karanovic, 2012)。
陰莖在不同的介形蟲之間差異甚大。即使在同一個屬的不同種之間,陰莖也有明顯差別。這種差異遠大於其它附肢的差異。這是因為,陰莖是用於交配的,而不同物種必須不能互相交配、具有生殖隔離。這種隔離機制主要是通過交配(包括前戲,即同一物種不同性別的識別,mate recognition)過程中同一物種雌雄個體能夠互相配合、完成交配,而不同物種之間不能來實現。一個簡單的例子,試想,陰莖的一部分要插入雌性外生殖器,因此其形狀、大小和所著生的位置,就要恰好與雌性陰道吻合。這種關係就好比“鑰匙和鎖”的關係。鎖不同,鑰匙也不同。
由於不同種的陰莖差別很大,因此它是十分重要的分類學特徵。常常僅根據陰莖就能輕鬆鑒定介形蟲。但是有些介形蟲類群只有雌性,進行孤雌生殖,因此只有通過對其它附肢的詳細觀察來定種。
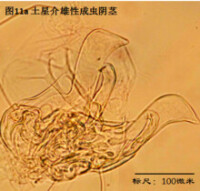
圖11a
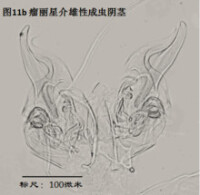
圖11b
圖12
4. 生態
介形蟲廣泛分佈於海洋、湖泊、池塘、沼澤、溪流、地下間隙水等各類水域中,極少數種類為陸生或半陸生。多為底棲種類,在底泥表面爬行、鑽入底泥淺層,或攀爬於水草之上。許多種類在第一和第二觸角上生有長的游泳毛,能遊動,但活動範圍仍多限於水底層附近,絕少來到水的中上層。因此,用浮游生物網很難捕到介形蟲。短背介這一類除外;它們介殼的腹部是平的,能倒懸並緊貼於水面之下。
食性雜,取食各種有機碎屑。但也許不同種或多或少有食性偏好吧。
有些種類可寄生於兩棲類和魚類;有寄生在人類肛門的報道。
對水溫、水的化學成分(離子組成、含氧量等)都有一定要求,不同屬種的適應範圍也不一樣。
5. 研究歷史
早在1748年,生物分類學的奠基人、瑞典自然學者Carolus Linnaeus,在Systema Naturae一書中,就對介形蟲動物進行了初次描述(Tressler, 1959)。但Linnaeus未對所描述的標本進行正式命名,後人只是根據他的描述,得知該標本屬於現在所稱的介形蟲。
1776年,丹麥人Otto Friedrich Müller描述了一些小型甲殼動物,將其歸入一個新設立的屬Cypris(金星介屬,Genus Cypris)中,分別命名為Cypris pubera,Cypris candida(現稱Candona candida)和Cypris laevis(現稱Cyclocypris laevis),等等。這就是最早被正式命名的介形蟲。隨後,在1785年,Müller在甲殼綱下建立了Entomostraca亞綱(Subclass Entomostraca),將Cypris屬和另一個新設立的屬Cythere(浪花介屬,Genus Cythere),以及其它一些並非介形蟲的甲殼動物歸入該亞綱(Baird, 1843; Hanai et al., 1985; 禹娜等,2004)。
1806年,著名的昆蟲學家、被譽為“昆蟲學王子”(Prince of Entomology)的法國人Pierre André Latreille,在他流放期間,建立了Ostrachoda亞綱(Subclass Ostrachoda)(也有說法認為Ostracoda亞綱是Latrielle於1802年建立的),並將介形類和枝角類從Entomostraca亞綱移入Ostrachoda亞綱。“Ostrachoda”後來被改寫為“Ostracoda”,成為現在所使用的“介形蟲”一詞。英文的ostracod和ostracode,都是從拉丁文Ostracoda派生而來(Hanai et al., 1985)。1821年,H.E. Straus將枝角類從Ostracoda亞綱中分出,從而使Ostracoda成為獨立的一類,即現在所稱的“介形蟲”。
繼Latrielle和Straus之後,越來越多新的介形蟲被發現並命名。隨著資料的積累,人們對介形蟲的軟體和外殼結構進行更加系統和詳細的描述。現生介形蟲的分類系統,在十九世紀晚期和二十世紀上半葉之間逐漸完善。在二十世紀,一些學者還對介形蟲的內部解剖結構和個體發育進行研究(Snodgrass, 1956)。與此同時,形態學和分類學的資料也在不斷積累(Tressler, 1959)。到了二十世紀後半葉,有人提出,生物的遺傳物質能記錄其進化歷程。由於介形蟲是現存的歷史最為悠久的生物類群之一,其DNA、RNA所包含的生物演化信息必然極為豐富。因此,一些學者開始通過研究細胞中的DNA、RNA來追溯介形蟲及其所代表的節肢動物的演化歷史和內在規律(Bode et al., 2010)。另一方面,介形蟲作為廣泛分佈的一類小型動物,它們在生態環境測評和保護研究方面的作用也漸漸為人們所認識(Pieri et al., 2012)。
至於對介形蟲化石的研究歷史,則追溯到1813年。當時,“介形蟲微體古生物學之父”(Father of Ostracode Micropalaeontology)、法國的Anselme Gaetan Desmarest從法國中部Allier地區的漸新世地層中發現了第一枚介形蟲化石標本,命名為Cypris faba(Hanai et al., 1985; Matzke-Karasz et al., 2007)。隨後,Jones(1850)在英格蘭南部更新世地層中進行了最早的對第四紀介形蟲化石的研究。此後,在十九世紀下半葉中,Jones,Prestwick,Sherborn,Chapman,Kirkby等人對多地的化石介形蟲進行了描述,積累了大量形態學和地層學資料,由此逐步建立了化石介形蟲的分類系統。
在二十世紀的大多數時間裡,介形蟲化石的研究著重於它們在地層中的分佈、尤其是在某一個地區的地層序列中的分佈,目的主要是對含化石地層進行劃分對比和確定地層時代。在這一時期,由於石油地質勘查和地質礦產調查的需要,歐、美、前蘇聯和中國在內的世界各國,都開始利用介形蟲和其它微體化石進行生物地層學的研究。在我國,介形蟲生物地層學的開拓者和奠基人是侯佑堂(1919–2010)。二十世紀中後期至今,在對介形蟲生物地層學深入研究的同時,古環境研究學者開始根據地層中化石屬種組成和殼體地球化學的變化,推測地質時期的環境波動。
如上所述,對現生介形蟲的研究和對介形蟲化石的研究是有區別的。對現生介形蟲的研究,主要是依靠它的“肉”,也就是介殼裡的那個身體——從水裡活捉介形蟲,依靠附肢的形態、剛毛的數目、位置和長短等來進行鑒定和分類,並提取DNA進行研究;儘管介殼的形態也具有一定分類學價值。對化石介形蟲的研究則幾乎僅僅圍繞介殼進行——從沉積物和地層里挖出介殼,研究不同種類的地層和時代分佈,從而對地層進行對比、劃分;研究特定時代介形蟲的種類組成、介殼化學成分,從而推測古代環境狀況。因此,現生介形蟲學家和化石介形蟲學家通常是不打交道的,儘管他們在某些有交叉的論題上合作。例如研究介形蟲的演化時,地層中的演化譜系可以與根據現生種類親緣關係得到的演化關係進行對比。再如,對某些現在還活著的介形蟲,當它們的介殼從沉積物中被找出來的時候,對它們的屬種鑒定就需要與現生種類的分類系統相互對照,統一到後者上來。Limnocythere inopinata具有不同瘤飾的各個表型,在很長時間內就曾被國內的化石介形蟲學者分為不同的物種;但通過對現生種的附肢的觀察發現,它們實際上是同一個東西(Yin et al., 1999)。最後,當古環境學家為地層中某種介形蟲所反映的環境是怎樣的而苦惱時,他們也需要請教現生介形蟲學家,這個種現在生活在什麼樣的環境中。
就現生介形蟲來說,在當前學術界,研究的中心在歐洲,而中國的研究遠遠落後。就化石介形蟲的研究而言,我國可以說曾經一枝獨秀,但或將日趨式微。
6. 應用
不同應用的羅列順序和篇幅未必反映其重要性。
1)地層對比和劃分。請見上文。
2)古環境研究。請見上文。
3)生態環境測評,以及生物多樣性保護
人們利用介形蟲的種類組成和物種多樣性來反映水域的水質和生態健康狀況(Pieri et al., 2012)。
對生態健康狀況的測評,原理是這樣的。在眾多介形蟲中,有世界廣布的屬種,也有地方性特有的屬種。廣布種適應性強,地方種通常只能適應某個地區的穩定的、原生態的環境。當某個地區的自然環境遭受破壞(通常是人為的)時,原生的生物群落就將被瓦解,而一些別的地區的物種,通常是那些適應能力強的廣布種就侵入並佔據了這個地方。這個過程中,生物群落的成分改變了,並且多樣性通常下降,因為被破壞、被污染的環境中只有少數適應能力較強的物種能夠生存。而當人們採取一些保護措施、恢復環境的水質、減少人類活動的侵擾,從而使環境逐漸好轉時,這個地區的生物多樣性就又會增加,一些原本瀕臨消失的地方性種又重獲新生,並且更多來自其它地方的、適應能力並不是那麼強的物種也加入到新的群落中來。人們注意到,這個次生的生物群落的組成,和人類來到之前的那個原生態的群落的組成是有所不同的,因為在破壞——恢復的過程中,某些土著物種被不可逆地淘汰掉了,而一些外來的移民則在這裡紮根。儘管成分變了,生物多樣性的增加仍然反映了環境保護措施的成效、生態健康狀況的改善。另一方面,同樣的生物多樣性,那些地方性屬種豐富的生境,就代表保護得很好、從未或很少遭受人類破壞的原生態狀況;而那些充斥著各種廣布種的生境,就很可能是曾經遭受人類活動破壞、之後逐漸修復的生境。
介形蟲當然不是唯一的能用於生態測評的物種(Boets et al., 2011)。但筆者認為,介形蟲、硅藻等小型生物,由於其分佈廣、繁殖快、數量多,因而與較大的生物相比,具有容易取樣、響應靈敏、更易滿足統計要求等優點。
利用生物群落對水質進行評價,是因為人們在各種水質化學指標之外,還希望看到一些直觀的、利用水生生物的生存狀況來反映水源好壞的指標。畢竟,新型污染物太多了,光看儀器測量有點不放心,總想看看裡面的魚蝦活得怎麼樣。而且從心理上,現在的人們也傾向於喝那些“原生態”的水。但是利用生物來進行水質評價,取代不了器測,充其量不過是一種補充手段。
介形蟲
介形蟲