生殖隔離
不能產生後代等的隔離機制
生殖隔離指由於各方面的原因,使親緣關係接近的類群之間在自然條件下不交配,或者即使能交配也不能產生後代或不能產生可育性後代的隔離機制,若隔離發生在受精以前,就稱為受精前的生殖隔離。
核, 論形態差, 殖隔離( isolation)應該是兩個群體能否真正分化成不同物種的關鍵, 這種隔離機制可以是地理的、行為的或其他方式; 而生殖隔離總會伴隨著一些形態或遺傳上的變化, 雖然這些特徵可能與生殖隔離本身並無多大關係, 但往往成為分類學家或分子進化生物學家區分種的依據。
殖隔離致隔離,地理隔離既有聯繫,也有區別。地理隔離由外因引起,而生殖隔離一般是在具有遺傳差異(內因)的基礎上才有可能(遺傳隔離)。同一地點的種、亞種的隔離完全是生殖隔離,但不同地點的種、亞種在許多情況下,似乎也會很快發生這種作用。關於生殖隔離,許多人進行過討論。斯特賓斯(G.L.Stebbins,1966)作了如下的分類:(1)合子形成前的機制。(i)生態隔離:雖在同一地域,但生態地位不同。(ii)季節隔離:開花期有分離。(iii)行為性隔離(ethological isola-tion):雌雄的性習性不同。(iv)機械隔離:基於生殖器官構造的差異。(v)配子不親和現象(gameteincompatibility):一方的配子體在另一方的生殖器官內死亡。(2)合子形成后的機制。(i)雜種死滅或弱勢(hybrid inviability,hybrid weakness)。(ii)雜種不育。(iii)雜種衰敗(hybrid break-down):F2以後死滅或弱勢。在自然界中這些因子是重複地起著隔離作用的。
生殖隔離地理隔離
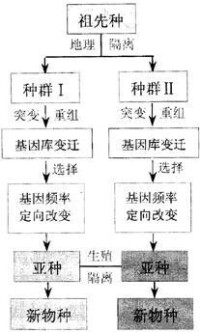
現代進化論認為,物種形成大致要經過3個階段:1)由於基因突變、染色體變異和基因重組,使種群中產生可遺傳的變異;2)自然選擇等因素作用於可遺傳的變異,使種群的遺傳結構(基因型頻率和基因頻率)發生了適應性的改變;3)不同種群由於地理隔離和生態隔離而加深了性狀分歧,逐漸形成亞種,一旦出現了生殖隔離,亞種就變成了新種。此外,在植物界異源多倍體的產生亦可導致新種形成。
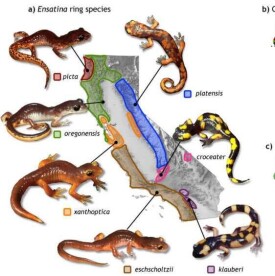
同一種生物由於地理上的障礙而分成不同的種群,使種群間不能發生基因交流的現象,叫做地理隔離。
由於食性、生活習性和棲息地點的不同,使幾個親緣關係接近的類群之間交配不易成功的隔離機制。例如有人報道5種近緣雀在樹冠的不同層次捕食不同的昆蟲,以及營巢的位置和交配產卵的繁殖季節不同,所以它們雖然生活在同一片森林,甚至同一棵樹上,也都因各有自己的生態位置而形成生殖隔離。
又稱時間隔離,新種形成過程中,合子形成前的隔離機制之一。生物一般都有一定的生育季節和時間,如動物的發情期、交配期;植物的開花期和授粉期等,如果同種群體間的生育季節和時間不同,就會造成季節隔離或時間隔離,阻止了基因的交流,從而導致生殖隔離的形成。例如,北美有3種蛙,同生在一個池塘里,但由於繁殖季節不同而構成了生殖隔離。林蛙在水溫達到7 ℃時,首先產卵;其次是笛蛙,在水溫達到12 ℃時開始產卵;鼓蛙在水溫達到16 ℃時才開始產卵。
植物傳粉后或動物交配后,由於生理上的不協調而不能完成受精作用的現象。例如,異種花粉落在柱頭上通常不能形成花粉管;有的即使形成花粉管也不能成功地通過花柱、到達胚珠。又如綠果蠅的精子在美洲果蠅的受精囊內很快就喪失遊動能力,而同種精子則可較長時間保持活動能力。
主要指由於植物花的結構與傳粉者形態結構不協調而造成的交配受精障礙。例如,大紅吊鐘柳,花紅色,有細長的管狀花冠;山吊鐘柳,花藍色,有較大的鐘狀花冠。二者在人工授粉時能形成雜種,但在自然條件下,前者以其紅色光和紅外光招引蜂鳥,管狀的花冠對蜂鳥細長的喙也相適應;後者以藍紫光與紫外光招引蜂類,鍾狀花冠也適於蜂類采蜜,但二者之間不能相互授粉。這樣,它們的花色和結構,便構成了物種間的隔離而阻止了基因的交流,此即為形態隔離。
由於交配行為不同,而使兩個或幾個親緣關係相近的類群之間交配不易成功的隔離機制。例如,紅蟋蟀、賓州蟋蟀和富爾頓蟋蟀在形態上無區別,但鳴聲的強弱和頻率不同。試驗證明,交配僅在發生同一鳴聲的種內進行,不同鳴聲的蟋蟀不進行交配。
一種雜交后的生殖隔離,不同種生物受精后,雖能形成合子,但合子不能存活;或合子雖能存活,但胚胎髮育到一定階段便死亡;或者胚胎髮育雖然完成,併發育為幼體,但不能生活到性成熟,故留下後代的概率很低。例如,以奧地利亞麻(♀)與多年生亞麻(♂)雜交,雜種的胚不能充分發育,因為胚和胚乳不協調。反交時,胚和胚乳雖較協調,雜種胚可以充分發育,但不能穿破母本組織形成的種皮,種子不能萌發,雜種也不能成活。
一種雜交后的生殖隔離。不同種生物雜交后雖能形成合子,併發育為成熟的F1雜種,但雜種不能產生後代。例如,馬和驢雜交所生的騾就不能生育。因為馬的染色體為64,驢的染色體數為62,騾的染色體數為63。馬的32對染色體中有18對端著絲粒染色體,驢的31對染色體中只有11對端著絲粒染色體,騾在形成生殖細胞時,染色體不能正常配對(即聯會紊亂)而不能完成減數分裂和形成正常的配子,故不能生育。 。不過野豬在全世界有27個亞種。亞種間和亞種內核型都有一些差異,染色體數(2n)在36-38之間,如西歐野豬2n=36或37,日本野豬2n=38,沒有繁殖障礙。普氏野馬(Przewalski's horse) 66個染色體,家馬(64個染色體)可以雜交生下可育的後代。野豬在全世界有27個亞種。彼此間沒有繁殖障礙,染色體數56的野牛和60對黃牛雜交有可育後代,獨龍牛、黃牛(Bos taurus)和野牛染色體數目(2n)分別為58、60和56條,也都有可育後代。盤羊屬, 染色體進化是從2n = 58到2n = 52。家羊(2n = 54),雜交有可育後代,藏野驢其實是2n=51~52。波斯野驢和土庫曼野驢都是2n=54~56,家驢有62條染色體。也有可育後代
也叫雜種破落,是一種雜交后的生殖隔離機制。種間雜交雖能形成能育的F1,但在F2或回交後代中,絕大部分重組基因型不相協調,無適應能力,不能存活。例如,有人以陸地棉的斑點變種為母本,與夏威夷棉雜交,F1能活也能育。但是F1植株上結出的106粒F2種子中,7粒胚小,不能發芽;32粒胚看來正常,但也不能萌發;9粒能穿破種皮,但子葉不能展開;22粒能長成幼苗,但不久即死亡;16粒長成弱苗,僅活了3周;20粒長成的幼苗,能活3周以上,但以後發育也不正常。這表明在F2的重組基因型中,存在著雜種衰敗機制,它們經不起強大的自然選擇壓力,最終也不能存活下來。
以上各種類型的生殖隔離,實質上都是阻礙不同物種間的基因交流,使各個隔離種群各有較強的穩定性,以保證在自然條件下各按著與環境相適應的方向發展。因此,物種是生殖隔離的種群,由地理隔離發展到生殖隔離是大多數物種形成的基本因素。
遠緣雜交的實質是打破物種間的生殖隔離,實現不同物種間的優良基因重組,創造為人類所需的植物新類型和新種質。物種間的生殖隔離是由不同的繁殖隔離機制來保持的,繁殖隔離機制阻止種間的基因流動,是防止生物種間雜交的生物學特性。根據隔離發生的主要時期將繁殖隔離機制劃分為兩大類:受精前的生殖隔離和受精后的生殖隔離。若隔離發生在受精以前,就稱為受精前的生殖隔離,其中包括地理隔離、生態隔離、季節隔離、生理隔離、形態隔離和行為隔離等;若隔離發生在受精以後,就稱為受精后的生殖隔離,其中包括雜種不活、雜種不育和雜種衰敗等。