反義RNA
能封閉基因表達的RNA
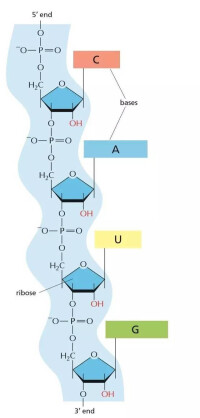
反義RNA是指與mRNA互補后,能抑制與疾病發生直接相關基因的表達的RNA。它封閉基因表達,具有特異性強、操作簡單的特點,可用來治療由基因突變或過度表達導致的疾病和嚴重感染性疾病。根據反義RNA的作用機制可將其分為3類:Ⅰ類反義RNA直接作用於靶mRNA的SD序列和(或)部分編碼區,直接抑制翻譯,或與靶mRNA結合形成雙鏈RNA,從而易被RNA酶Ⅲ降解;Ⅱ類反義RNA與mRNA的非編碼區結合,引起mRNA構象變化,抑制翻譯;Ⅲ類反義RNA則直接抑制靶mRNA的轉錄。
反義RNA
細胞中反義RNA的來源有兩種途徑:第一是反向轉錄的產物,在多數情況下,反義RNA是特定靶基因互補鏈反向轉錄產物,即產生mRNA和反義RNA的DNA是同一區段的互補鏈。第二種來源是不同基因產物,如OMPF基因是大腸桿菌的膜蛋白基因,與透性有關,其反義基因MICFZE則為另一基因。
反義RNA的分類和作用機制:下表總結了原核細胞內天然存在的11種反義RNA。這些反義RNA按其作用機制可經分為三大類。
調節水平反義RNA靶RNA分類功能來源
轉錄后水平micFRNAompFmRNA1AOmpF合成染色體
oopRNAcⅡmRNA1B溶菌-溶源噬菌體
sarRNAantmRNA1A溶菌-溶源噬菌體
ouTRNA轉位酶mRNAⅠA轉位作用轉位子
finpRNAtraJmRNAⅠADNA轉位
sokRNAhokmRNAⅠA殺死作用
copARNArepAmRNAⅡ複製質粒
R1162RNArepⅠmRNAⅡ複製
pT181RNArepCmRNAⅡ複製
轉錄水平ticRNACAPmRNAⅢcAMP結合蛋白染色體
複製前水平RNAⅠRNAⅡⅢDNA複製質粒
Ⅰ類:這類反義RNA直接作用於其靶mRNA的SD序列和/或編碼區,引起翻譯的直接抑制(ⅠA類)或與靶mRNA結合后引起該雙鏈RNA分子對RNA酶Ⅲ的敏感性增加,使其降解(ⅠB類)。Ⅱ類:這類反義RNA與mRNA的SD序列的上游非編碼區結合,從而抑制靶mRNA的翻譯功能。其作用機制尚不完全清楚,可能是反義RNA與靶mRNA的上游序列結合後會引起核糖體結合位點區域的二級結構發生改變,因而阻止了核糖體的結合。Ⅲ類:這類反義RNA可直接抑制靶mRNA的轉錄。
ticRNA(transcriptioninhibitorycomplementaryRNA)是大腸桿菌中CAP蛋白(cAMP結合蛋白)的mRNA的反義RNA。ticRNA的基因的啟動子可被cAMP-CAP複合物所激活,從CAPmRNA的轉錄起始位點上游3個核苷酸處開始,以CAPmRNA的模板DNA鏈的互補鏈為模板,合成ticRNA。ticRNA具體長度不清楚,但是它是5'端一段正好和CAPmRNA的5'端有不完全的互補,可以形成雙鏈的RNA雜交體。而在CAPmRNA上緊隨雜交區之後的是一段約長11bp的A,U豐富區。這樣的結構十分類似於ρ不依賴性的轉錄終止子的結構,從而CAPmRNA的轉錄剛剛開始不久后即迅速終止。從這個例子中我們可以看到CAP蛋白合成的自我調節作用。當CAP合成達一定量后,即可與cAMP結合成cAMP-CAP複合物。再激活ticRNA的啟動子轉錄出ticRNA,反過來抑制CAP-mRNA的合成。
在原核生物中反義RNA具有多種功能,例如調控質粒的複製及其接合轉移,抑制某些轉位因子的轉位,對某些噬菌體溶菌-溶源狀態的控制等。下文僅舉數例。
調控細菌基因的表達
反義RNA對編碼CAP的基因的調控作用已如前述。這裡再介紹一下micFRNA對ompF基因的表達的調控。ompF蛋白質是大腸桿菌的外膜蛋白的主要成分之一。micFRNA是從另一基因(ompC基因)附近的DNA序列轉錄而來,和o-mpFnRNA的5'端有70%的序列互補,因此在體外micFRNA可以抑制ompFmRNA的翻譯。但是這種抑制作用在體內是否重要尚有疑問,因為缺失micF基因的菌株其ompF蛋白的表達只受到輕微的影響。
噬菌體溶菌/溶源狀態的控制
反義RNA也參與了λ和P22噬菌體的溶菌/溶源狀態的控制。P22噬菌體編碼一種抗阻遏蛋白Ant,它可以抑制許多λ樣噬菌體的阻遏蛋白與DNA的結合。這對於剛剛感染細胞的P22建立λ樣原噬菌體(prophage)是有益的。但是Ant必須在嚴格的控制下,否則Ant的過分表達必將阻止溶源狀態的建立,而成為溶菌性的噬菌體。Ant蛋白質表達的控制是利用反義RNA(sarRNA)能與antmRNA的翻譯起源區互補結合,從而抑制antmRNA翻譯成Ant蛋白。
在λ噬菌體中cⅡ蛋白控制著溶菌或溶源狀態的選擇。cⅡ蛋白可以激活λPre啟動子,該啟動子控制的基因是λ噬菌體整合作用所必須的,且同時能抑制λ噬菌體的複製。cⅡ蛋白的另一功能是延緩λ晚期基因的表達,其作用機制是cⅡ蛋白激活PaQ的啟動子,轉錄出PaQRNA。PaQRA是編碼Q蛋白的mRNA的反義RNA。因此,PaQRNA能與QmRNA配對雜交而抑制其翻譯,而Q蛋白早已知道是晚期基因表達的激活蛋白。
cⅡ基因本身的表達還受到稱為oopRNA的反義RNA的調控。oopRNA與cⅡ基因的3'端互補,但其具體作用機制尚不清楚。
IS10轉位作用的抑制
outRNA是一種反義RNA,可以和IS10編碼的轉位酶mRNA(INRNA)5'端結合而抑制其翻譯,當細胞內只有一個考貝IS10時,只能生成很少量的outRNA,故轉位酶仍可生成。但當IS10的考貝數增多時,outRNA愈來愈多,其控制作用亦明顯增強,所以稱為多考貝抑制現象。這種現象可以防止IS10的過量堆積引起的細胞損害。
既然反義RNA在原核生物中對基因表達起著重要的調控作用,那麼人工設計在天然狀態下不存在的反義RNA來調節靶基因的表達,想必也是可能的。這已在不少實驗中得到證實。
1.由於目前對靶mRNA的SD序列的上游區的結構了解甚少,因此,在要設計Ⅱ類反義RNA用於和靶mRNASD序列上游區結合,以期達到調節該mRNA翻譯的目的是比較困難的。
2.Ⅲ類反義RNA是和mRNA的起始處結合而形成類似ρ-不依賴性的轉錄終止子而使轉錄水平上抑制靶基因的表達。因此,要設法在靶mRNA上找到一段連續的U序列,就可以設計出反義RNA,與該U序列上游的mRNA鏈互補,以形成ρ-不依賴性終止子。理想的作用位點是在靶mRNA的5'端上游的非編碼區,以免受核糖體的影響。
3.只要靶基因的核苷酸順序已經知道,就可以人工設計出Ⅰ類反義RNA。有時還可設計同時具有Ⅰ類和Ⅲ類反義RNA功能的反義RNA。
我們還可以設計出天然存在的反義RNA的反義RNA來。這樣就可以拮抗原始反義RNA對靶mRNA的抑制作用。而達到激活或加強某個靶基因的表達的目的。然而並不是所有的mRNA對其相應的Ⅰ類反義RNA都敏感。例如,有的mRNA壽命很短,只有1-2分鐘,它們和反義RNA結合的機會較少,因而就較不敏感。反之,另一些mRNA則很穩定,壽命可達十多分種,則其對相應的反義RNA的抑制作用就很敏感。此外,反義RNA本身的穩定性有很大的實際意義。顯然,穩定的反義RNA對靶mRNA的調節作用比不穩定的反義RNA要好。
隨著分子生物學和遺傳工程的發展,基因治療應運而生,反義技術是其中一種,它的基礎是根據核酸雜交原理設計針對特定靶序列的反義核酸,從而抑制特定基因的表達,包括反義RNA、反義DNA及核酶(Ribozyme),它們通過人工合成和生物合成獲得。
反義DNA是指一段能與特定的DNA或RNA以鹼基互補配對的方式結合,並阻止其轉錄和翻譯的短核酸片段,主要指反義寡核苷酸,因更具藥用價值而倍受重視。
核酶(ribozyme)是具有酶活性的RNA,主要參加RNA的加工與成熟。天然核酶可分為四類:(1)異體催化剪切型,如RNaseP;(2)自體催化的剪切型,如植物類病毒、擬病毒和衛星RNA;(3)第一組內含子自我剪接型,如四膜蟲大核26SrRNA;(4)第二組內含子自我剪接型。利用反義技術研製的藥物稱反義藥物。反義藥物作用於產生蛋白的基因,因此可廣泛應用於多種疾病的治療,如傳染病、炎症、心血管疾病及腫瘤等。與傳統藥物比較反義藥物更具選擇性及效率,因此也更高效低毒。基於上述特點反義藥物已成為藥物研究和開發的熱點。而且反義技術還可以應用於生物科學的基礎研究。
長的反義RNA並不一定比短的反義RNA更為有效;
在原核生物中針對SD序列及其附近區域的反義RNA可能更有效;
在真核生物中,對應於5'端非編碼區的反義RNA可能比針對編碼區的反義RNA更有效;
盡量避免在反義RNA分子中出現自我互補的二級結構;
設計的反義RNA分子中不應有AUG或開放讀框,否則該反義RNA亦會與核糖體結合而影響其與靶mRNA的配對結合;
進一步還可以將帶有ribozyme結構的RNA連在反義RNA的3'端尾上,當反義RNA與靶mRNA雜交后,即可利用其酶活性來降解靶mRNA。
此外,為了增強反義RNA的作用,還可以採取一些額外措施,
例如:由於反義RNA對靶mRNA的抑制作用有劑量依賴性,所以在構建反義RNA基因時,要選擇強的、可以誘導的啟動子以增強反義RNA本身的表達;
構建許多個反義RNA基因串連在一起,以得到線性重複的多拷貝基因,對提高反義RNA的表達也有利;RNA酶Ⅲ可以降解RNA:RNA雜交體,所以在構建反義RNA基因時,可將RNA酶Ⅲ的基因也同時轉化到靶細胞中並進行表達。這樣,當反義RNA與靶mRNA結合后,RNA酶Ⅲ即可將其降解。這顯然有利於反義RNA的抑制作用;
近年來,有關反義RNA的研究進展迅速,已經應用到抗病毒感染,研究癌基因的作用機制,探索腫瘤治療的可行途徑等方面。在今後一段時間內,有關反義RNA的研究肯定將會有更加迅速的進展和更廣闊的應用前景。
反義RNA3'端帶有莖環結構或類似ρ-不依賴性終止子結構時,可以穩定RNA分子;
Gorski等還發現在T4噬菌體的基因32mRNA的5'端上游的莖環結構及其附近的序列亦可穩定RNA分子。所以在設計反義RNA基因時,最好將產生3'及5'端這種二級結構的序列克隆在反義RNA基因的兩端。Hirashima等1986年發現,針對靶mRNA的SD序列和AUG區域的反義RNA,要比單純對編碼區域的反義RNA更為有效。1989年Hirashima又發現,針對SD序列和它的上游區域(但不包括AUG)的反義RNA更為有效。在真核生物中,針對5'端非編碼區的反義RNA更有效。但也有實驗表明針對第一內含子的反義RNA也同樣有效。
基本信息
- 中文名
- 反義RNA
- 外文名
- antisense RNA
- 概念
- 與mRNA互補的RNA分子
- 作用
- 抑制mRNA的翻譯
- 來源
- 反向轉錄的產物、不同基因產物
- 分類
- 三類