進化趨勢
進化趨勢
進化趨勢指在相對較長的時間尺度上,一個線系或一個單源群的成員表型進化改變的趨向。生命進化的總體趨勢是體型的多樣化和複雜化、遺傳信息的複雜化、體積趨於增大以及脊椎動物腦趨向於增大。當然,有時也會出現簡化或退化。
地球上的生命呈現出一種永不停息的進化趨勢,即它們絕不放過任何微小的變異(這裡不管是何種機制)機會,只要能帶來改進或革新,能適應未被完全佔用的生境或能獲得競爭能力去侵佔已被其它物種佔用著的生境,它就會延續和發展下去,形成新的變型,甚至分化出新的物種。進化也是一個頗具爭議的辭彙,人們往往意指進步,在這裡的進化廣義地指演化。
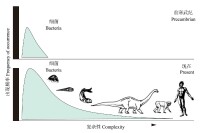
圖1 複雜性的進化
需要指出的是,生命系統的複雜化趨勢,並不意味簡單生命的消失,事實上,簡單的生命(如古老的細菌)仍然無處不在,也佔據著屬於它們的那一份生態位,也仍然未停止繼續分化。說白了,有些功能(如生物殘體的分解)並不適合複雜的生命去干,細菌仍然握有絕活,不可替代。因此,從某種意義上來說,應該說是進化伴隨著生命的多樣化(包括複雜化)的發展,添加了許多複雜化(一些人喜歡叫進步)的類群。也許還有一些生命保持了變化不大的狀態(但絕對沒有一點變化也是不可能的),有些甚至出現“退化”(結構簡化)。因此,進化的結果絕非只有普遍“進步”,但同時也不能完全否認“進步”的存在。從本質上來看,由於生命演化中普遍存在的複雜化現象,自然選擇推動的物種分化註定是一種不可逆過程。
另一個不可否認的事實是,在生命演化過程中也出現了大量的結構簡化的例子。複雜性實質性減少普遍出現於寄生蟲的生活模式,涉及到成千上萬的種類,它們寄生於宿主體內吸食宿主血液或宿主已消化的食物,已不再需要消化或運動器官,但為了特殊需要,可能會有一兩種新的器官出現—抓住宿主的鉤、吸食食物的吸管等,但它們幾乎變成了生殖系統構成的袋子或管子—簡單的生殖機器,一種依附在宿主的內部器官罷了。這樣,這些寄生蟲損失的器官比增加的多得多,所以結構大大地簡化了(古爾德2009)。從天南星科的大型祖先植物演化出最小的被子植物—漂浮在水上的蕪萍是另一個結構簡化的很好例子。
因此,簡單化也是進化的方向之一是毋庸置疑的,這一方面從基因的隨機突變(如基因的缺失)的概率上來講是可能的,另一方面一些複雜的生命由於生存環境的變遷(如氣候乾旱化,從自由生活轉為寄生,等等)也能向簡單化的方向發展,如上所說的寄生蟲的結構一般都會比其祖先簡單。
事實上,生命演化的過程不僅僅是進化(通常是結構複雜化),也包括退化(通常是結構簡化),還有一些類群變化不大的類群。因此,生命演化的結果不一定都是“進步”,也有“退步”。實際上所謂“進步”也是相對的,離不開生存條件。在一定條件下的複雜“進步”當巨大的環境災變降臨時,也會遭遇滅頂之災(如恐龍)。因此,傾向於給進步加上引號。
一些物種趨於簡單化的理由似乎容易理解,但是為何絕大多數生命要向複雜的方向進化?為何簡單原始的生命(細菌)還能一如既往在屬於它們的“領地”中生存至今?生命複雜化是進化的原因(進化生物學中的進步論學派)還是結果?
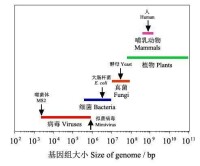
圖2 各種生物的基因組大小比較
即便如此,從趨勢上來看,隨著生命的進化,生物複雜性不斷增加,儲存遺傳信息的基因組也不斷增大,也驅動生物體積的逐漸增大。
早在一個多世紀以前,美國古生物學家柯普(Cope 1896)就觀察到化石記錄中常常出現體積增長的趨勢,被稱之為柯普定律(Cope’s law)。譬如,馬是從和狗一般大小的動物持續進化,愈變愈大的,同時腳趾持續萎縮,終於變成蹄;象的始祖本來只有豬那麼大,歷經數千萬年的進化,後代終於變成龐然巨物,同時象牙逐漸伸長,愈形複雜(齊默2011)。

圖3 從猴→猿→人演化過程中體積的變化趨勢
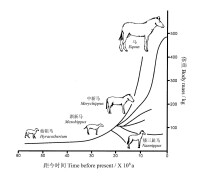
圖4 北美化石馬體積進化格局
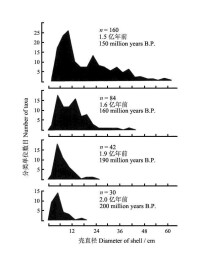
圖5 進化時期菊石體積頻數分佈的擴展
柯普現象雖然普遍存在,但是也會出現許多例外,有些類群可能體積變化不大,有些甚至可能會變小。此外,變大的趨勢也不是無限增加,大的物種也不一定具有永恆的優勢,也是局限於一定的地質歷史時期以及一定的氣候與生態環境背景,像巨大的恐龍的滅絕就是一個範例。無論如何,在過去的地質歷史時期,許多類群向體型變大的方向進化或者體型多樣化是不爭的事實。
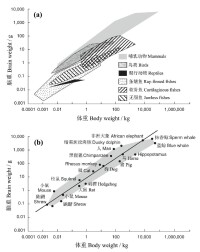
圖6 脊椎動物腦重和體重的關係(對數尺度)
人類腦容量的顯著增加是人類意識智能化的重要物質基礎。在600萬年前,我們祖先的大腦大約只有我們現在的1/3大小,有理由相信,他們的心智水平與其它猿相似,可能通過咕噥和手勢進行交流,不會用火或製造複雜的石制工具,也還無法深刻理解其他同伴的想法或感受,雖然這只是一種可能永遠都無法得到證實的猜測(齊默2011)。
基本信息
- 中文名
- 進化趨勢
- 外文名
- trend of evolution
- 應用學科
- 遺傳學、進化遺傳學