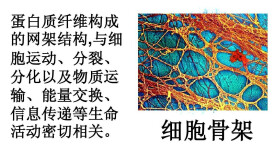
細胞骨架
細胞骨架
狹義的細胞骨架(cytoskeleton)概念是指真核細胞中的蛋白纖維網架體系(微管(microtubule, MT)、微絲(microfilament, MF )及中間纖維(intermediate filament, IF )組成的體系)。它所組成的結構體系稱為“細胞骨架系統”,與細胞內的遺傳系統、生物膜系統、並稱“細胞內的三大系統”。直到20世紀60年代后,採用戊二醛常溫固定,才逐漸認識到細胞骨架的客觀存在。是真核細胞藉以維持其基本形態的重要結構,被形象地稱為細胞骨架,它通常也被認為是廣義上細胞器的一種。廣義的細胞骨架概念是細胞核骨架、細胞質骨架、細胞膜骨架和胞外基質所形成的網路體系。核骨架、核纖層與中間纖維在結構上相互連接,貫穿於細胞核和細胞質的網架體系。
細胞骨架(cytoskeleton)是指真核細胞中的蛋白纖維網路結構。發現較晚,主要是因為一般電鏡制樣採用低溫(0-4℃)固定,而細胞骨架會在低溫下解聚。直到20世紀60年代后,採用戊二醛常溫固定,才逐漸認識到細胞骨架的客觀存在。真核細胞藉以維持其基本形態的重要結構,被形象地稱為細胞骨架,它通常也被認為是廣義上細胞器的一種。
細胞骨架不僅在維持細胞形態,承受外力、保持細胞內部結構的有序性方面起重要作用,而且還參與許多重要的生命活動,如:在細胞分裂中細胞骨架牽引染色體分離,在細胞物質運輸中,各類小泡和細胞器可沿著細胞骨架定向轉運;在肌肉細胞中,細胞骨架和它的結合蛋白組成動力系統;在白細胞(白血球)的遷移、精子的遊動、神經細胞軸突和樹突的伸展等方面都與細胞骨架有關。另外,在植物細胞中細胞骨架指導細胞壁的合成。
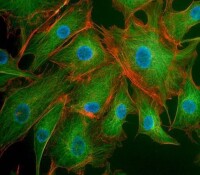
細胞骨架
從各種組織中提純微管蛋白可以發現還存在一些其他蛋白成分(5%-20%),稱之謂微管相關蛋白(microtube associated proteins MAPs)。這些蛋白具有組織特異性,表現出從相同αβ二聚體聚合形成的微管具有獨特的性質,已從人類不同組織中發現了多種α及β微管蛋白,並追蹤微管基因表現出部分基因家族,某些基因被認為是編碼獨特的微管蛋白。
微管形成的有些結構是比較穩定的,是由於微管結合蛋白的作用和酶修飾的原因。如神經細胞軸突、纖毛和鞭毛中的微管纖維。大多數微管纖維處於動態的聚合和災變(一種突然的,迅速的,一般不可逆轉的分解)狀態,這是實現其功能所必需的性質(如紡錘體)。與秋水仙素(colchicine)結合的微管蛋白可加合到微管上,並阻止其他微管蛋白單體繼續添加,進而破壞紡錘體的結構,長春花鹼具有類似的功能。紫杉酚(taxol),能促進微管的聚合,並使已形成的微管穩定,然而這種穩定性會破壞微管的正常功能。這些藥物可以利用破壞微管功能以阻止細胞分裂,成為癌症治療的新希望。
在人類至少發現兩種明顯區別的α-微管蛋白及三種明顯區別的β-微管基因,它們產生具有特定功能的微管蛋白mRNA,由於這些編碼在結構組分上十分近似蛋白質分子,在不同組織存在多少特異性的具有差異表達的微管蛋白亞型,尚待深入研究。
除了α-與β-微管蛋白有編碼相似的不同變異型,近幾年來又發現了多種編碼差異更大的新的微管蛋白,形成不同的基因家族。其中gamma微管蛋白位於細胞內的微管組織中心(microtubule organizing center, MTOC),是用以提供α及β微管蛋白進行聚合反應形成微管的起始核心。而delta與epsilon則被認為與中心體(centrosome)的結構與形成有關。其他尚有eta, zeta, theta等等多種變異,不過通常僅存在少數幾種真核單細胞生物如原蟲或纖毛蟲里,可能跟這些生物獨特的結構與生理習性有關,進一步詳情仍待研究。
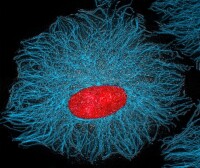
細胞骨架
微絲主要由肌動蛋白(actin)構成,和肌球蛋白(myosin,一種分子馬達蛋白)一起作用,使細胞運動。它們參與細胞的變形蟲運動、植物細胞的細胞質流動與肌肉細胞的收縮:
植物細胞的細胞質流動:微絲中的actin(肌動蛋白)與myosin(肌球蛋白)在細胞質形成三維的網路體系。actin位於外質,myosin位於內質。 myosin連結著細胞質顆粒,由ATP供給能量,myosin與細胞質顆粒的結合體沿著actin filament滑動,從而帶動整個細胞質的環流。
變形蟲運動(amoeboid movememt,阿米巴運動):肌肉細胞的收縮:
如同微管蛋白,肌動蛋白的基因組成一個超家族,並組成多種極為相似的結構。例如,各種肌肉細胞有不同的肌動蛋白:①骨骼肌的條紋纖維;②心肌的條紋纖維;③血管壁的平滑肌;④胃腸道壁的平滑肌。它們在氨基酸組分上有微小的差異(大約在400個氨基酸殘基序列中有4-6個變異),在肌肉與非肌細胞中都還存在β及γ肌動蛋白,它們與具有橫紋的α肌動蛋白可有25個氨基酸的差異。
G-肌動蛋白單體(含ATP)可聚合為呈纖維狀的F-肌動蛋白(含ADP),它們可由Mg2+及高濃度的K+或Na+誘導而聚合,聚合后ATP水解為ADP及C-肌動蛋白ADP單體,組成F-肌動蛋白。在骨骼肌的細肌絲(thin filament,由肌動蛋白構成)與粗肌絲(thick filament,由肌球蛋白構成)相互作用而使肌肉收縮(肌球蛋白可以起作肌動蛋白激活的ATPase的作用)。肌球蛋白也存在於哺乳動物的非肌細胞中(但以非聚合狀態存在)。
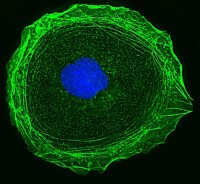
細胞骨架
以上這些結構單元並非是一成不變的,而是隨細胞的生命活動而呈現高度的動態性,它們均由單體蛋白以較弱的非共價鍵結合在一起,構成纖維型多聚體,很容易進行組裝和去組裝,這正是實現其功能所必需的特點。
長期以來,人們認為細胞骨架僅為真核生物所特有的結構,但近年來的研究發現它也存在於細菌等原核生物中。
目前為止,人們已經在細菌中發現的FtsZ、MreB 和CreS 依次與真核細胞骨架蛋白中的微管蛋白、肌動蛋白絲及中間絲類似。FtsZ 能在細胞分裂位點裝配形成Z 環結構,並通過該結構參與細胞分裂的調控;MreB能形成螺旋絲狀結構,其主要功能有維持細胞形態、調控染色體分離等;CreS存在於新月柄桿菌中,它在細胞凹面的細胞膜下面形成彎曲絲狀或螺旋絲狀結構,該結構對維持新月柄桿菌細胞的形態具有重要作用。
細胞在病理情況下常常會出現細胞骨架系統異常。如阿爾茨海默症患者,在腦神經元中發現有大量扭曲變形的微管和大量受損的中間纖維;在惡性轉化的細胞中,常表現為微管減少和解聚,細胞骨架異常可增強癌細胞的運動能力。研究表明,微絲束及其末端黏著斑的破壞以及肌動蛋白小體的出現,與腫瘤細胞的浸潤和轉移特性有關。
羊膜刺穿示意圖
不僅如此,細胞骨架還包含有很多結構單元的附屬蛋白質,比如:
分子馬達(molecular motors): 動力蛋白(dynein), kinesin, myosin
結合蛋白:vinculin, cofilin, tropomyosin等等
廣義的細胞骨架還包括核基質(nucleoskeleton)、核纖層(nuclear lamina)和細胞外基質(extracellular matrix),形成貫穿於細胞核、細胞質、細胞外的一體化網路結構。
細胞骨架(cytoskeleton)是指真核細胞中的蛋白纖維網路結構。發現較晚,主要是因為一般電鏡制樣採用低溫(0-4℃)固定,而細胞骨架會在低溫下解聚。直到20世紀60年代后,
細胞骨架
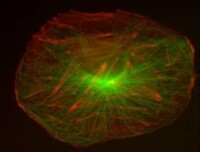
細胞骨架