光敏色素
光敏色素
吸收紅光-遠紅光可逆轉換的光受體(色素蛋白質),稱之為光敏色素(phytochrome)。
光敏色素分佈在植物各個器官中,黃化幼苗的光敏色素含量比綠色幼苗多20~100倍。禾本科植物的胚芽鞘尖端、黃化豌豆幼苗的彎鉤、各種植物的分生組織和根尖等部分的光敏色素含量較多。一般來說,蛋白質豐富的分生組織中含有較多的光敏色素。在細胞中,胞質溶膠和細胞核中都有光敏色素。
自1959 年發現光敏色素以來 ,人們就開始對其分子結構進行了研究。早期的研究發現 ,它是由線狀排列的四個吡咯環(生色團 chromophore)與一個蛋白質與硫醚鍵相連的色素蛋白複合體 ,而這個蛋白質是由兩個分子量在 1202127 KD的多肽聚合而成。而且 ,人們也通過研究發現 ,光敏色素都是以兩種不同的形式存在的 ,即紅光吸收型 Pr)和遠紅光吸收型( Pf r) ,Pf r 是生理激活型 ,其最大吸收峰在 730nm左右 ,而 Pr 是生理失活型 ,其最大吸收峰在 660nm左右 ,並且這兩種形式的光敏色素是可以相互轉變的 ,它們的轉變是生色團異構化的結果。近年來 ,人們利用免疫學、分子生物學及生物突變技術等研究表明 ,不同種類的光敏色素蛋白是由不同的基因編碼而成的。在擬南芥( A rabidopsis thal iana) 中,編碼光敏色素的蛋白基因有五種 ( phyA , phyB , phyC ,phyD , phy E)。被子植物的光敏色素基因是以家族形式存在的,其中對擬南芥的五種光敏色素基因研究得較為清楚。同時, Sharrock等也測定了擬南芥中 phyA ,phyB , phyC的cDNA順序。隨後 ,Clark 等又於 1994 年測定了后兩種 phyD 和 phy E 的 cDNA 順序。擬南芥的phyA 基因在黃化幼苗中含量非常豐富,但很容易被降解,而 phyB , phyC , phyD , phy E 的含量很少,並且對光很穩定。Prat t 等對番茄( L ycopersicon esculent um) 的phy 基因研究表明,其中也含有 5 種 phy 基因,其中的 4種與擬南芥對應相似,還有一種新的 phyF。Alba 的研究表明 ,番茄的 phyF 和擬南芥的 phyC 是同源的。因此,童哲曾提出, phy 基因與植物種類的進化有關。而且,Alba認為 ,不同種類的 phy 基因其進化速率也是不一樣的,其中 phyC 和 phy E 的進化速率是 phyA 和phyB 的1133 倍,這說明, phyA 和phyB 基因家族是比較保守的,同時, Kret sch利用突變技術的實驗也證明了這一點。最近 ,Park等利用生物物理和生物化學等手段發現 ,光敏色素包括有兩個中心區域 ,即 N 末端光感受區和 C末端光調節區 ,而且他還認為 ,其它作用因子是通過與光敏色素 C末端而發生作用的。
光敏色素的生理功能以前對光敏色素生理功能的研究大都集中在植物的外部形態和生理方面 ,如莖的伸長、葉的擴展、種子萌發、光周期誘導、小葉的偏下性運動等。近些年來 ,由於生物物理學、免疫學、分子生物學及生物定點突變技術在光敏色素研究中的廣泛應用 ,使人們越來越從生物化學和分子水平來闡述光敏色素的生理功能。
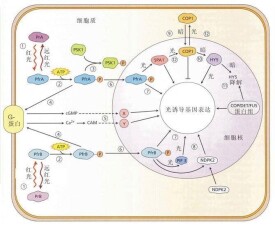
以前的研究發現 ,光對植物的下胚軸生長有抑制作用 ,但不同的植物其作用的光譜是不一樣的。對單子葉植物如水稻、燕麥等 ,短暫的紅光(RL)能抑制其下胚軸的生長 ,遠紅光(FR)有逆 轉作用 ,其反應是低輻照度反應(L FR) ,而對雙子葉植物 ,需要的是連續的 FR ,即光敏色素介導的高輻照度反應(HIR) 。Elzenga等用豌豆的葉表皮細胞進行研究 ,發現生色團突變株 Pcd2 和光敏色素B 突變株 ,其葉子在紅光下的大小明顯小於白光下的大小 ,而且他還發現 ,葉子的減小不是細胞分裂受抑制的結果 ,而是細胞伸長受到抑制的緣故 ,他提出 ,光是通過光敏色素影響細胞的酸化 ,進而影響其生長的。Quail等發現 ,黃花幼苗的抑制主要是 PhyA 介導的 FR2HIR反應 ,而在已轉綠的植物莖的生長抑制中是由 PhyB 介導的 RL2HIR反應。最近 ,Bert ram[10 ]對西紅柿再生芽的獲得進行了研究 ,發現再生芽至少是由PhyB1 和 PhyA兩種光敏色素所調節 ,而且 , PhyB1 是從下胚軸各個位置誘導再生芽 ,而 PhyA 卻沒有從下胚軸的中間和基部誘導再生芽。Bossen在研究 RL 誘導綠豆原生質體膨大過程中 ,發現 Pf r 會引起 G蛋白、Ca2 +、CAM、 PKC、 cAMP 酶的活化 ,說明這些分子都參與了信號傳導。但是 ,光敏色素對細胞生長的調控是非常複雜的 ,而且 ,同一種光敏色素分子在不同種類的植物中對細胞生長的調控可能是不同的 ,這一點還需要進一步研究。
自 1960 年 Marcuss 發現 3-2磷酸甘油醛脫氫酶(NADP)的活性受光的調節以來 ,人們通過大量的實驗來研究光與酶活性的關係。而且 ,此種方法也被證明是研究光敏色素作用機理的有效切入點。至今 ,人們已經發現植物體內有 60 多種酶或蛋白質受光的調控 ,這些酶涉及到植物的許多重要代謝途徑 ,如光合作用、呼吸作用、氮素同化、核酸的合成與降解等等。其中 ,紅光能提高硝酸還原酶活性在許多黃化植物中都已得到了驗證。最近 ,在擬南芥中發現 ,核苷二磷酸激酶 2 (NDPK2)是光敏色素信號傳遞的一個組分 ,它通過與 Pf r 結合而增強其活性 ,最終影響植物的光形態建成。但是 ,Sharma等卻認為 ,光敏色素能作為一種絲氨酸激酶使其自身磷酸化或使其它蛋白磷酸化 ,而後從細胞溶質轉到細胞核 ,在核內與轉錄因子相互作用而影響到基因的表達。
黃化幼苗照射 RL 後會去黃化 ,證實紅光促進葉綠素的合成。玉米只照射 1min 后 ,其葉綠體中 psaA、psaB、 atp E、 atpH、 psbO 等基因的轉錄物就增加 2~4 倍 ,照射 FR后 ,其 mRNA的增加卻大大減少。光和碳循環中的 Rubisco 大亞基編碼基因rbcL 和小亞基編碼基因rbcS的轉錄可以被光敏色素協調地調節。此外 ,光還能促進翻譯的起始過程 ,使葉綠體蛋白大大合成。這些都說明 ,光合作用和光形態建成是協調進行的 ,而且 ,光形態建成應該是光合作用的先決條件。Haupt (1965)曾研究紅光和遠紅光對轉板藻葉綠體運動的影響 ,發現紅光使它的葉綠體寬面朝向光的一面 ,而遠紅光則逆轉這個過程 ,說明是 Phy 介導的紅光/遠紅光效應。而 Sato的研究發現 ,Phy 介導的葉綠體的向光和避光運動是通過微管系統而進行的 ,這說明 ,Phy對葉綠體的調控是通過多種機制進行的。
1920年 , Garner 和 Allard 就發現了光周期現象 ,但是 ,直到 1959 年光敏色素被發現以後 ,人們才真正從內部生理和分子水平來解釋植物的光周期現象 ,而且 ,進一步的實驗證明 ,光周期不僅能影響到植物的開花 ,而且還能影響到色素的形成、葉子的脫落、種子的休眠等一系列生理過程 ,童哲曾證明光敏色素通過光周期參與調節光敏稻的雄性器官發育。Devlin 等利用突變技術 ,發現擬南芥的phyE缺失突變體的開花時間提前 ,而且 ,它也是 弱 了 PhyA、PhyB 幼 苗 對 遠 紅 光 的 反 應。Kret sch等發現 ,PhyB 的一個亞中心是高度保守的 ,這個亞中心的一個氨基酸發生了交換 ,這個突變株表現出對持續紅光的敏感性 ,在短日照條件下 ,其表型發生了改變。而最近 Childs 的研究也說明 , PhyB 基因中由於一個鹼基的缺失而導致 PhyB 功能的喪失 ,引起植物對光周期的敏感性降低。擬南芥的光敏色素生色團突變體hy1和 hy2 在長日和短日條件下其開花都比野生型提前。最近 ,Devlin的研究表明 ,擬南芥的 PhyB 突變體其開花提早 ,而且他還認為 ,PhyD 能通過控制開花時間來控制植物的避陰反應。在植物的光周期反應中 ,至少涉及到 L FR 和 HIR兩種類型的光敏色素作用。而且 ,一般性的結論是:PhyA與長日植物(LDP)誘導的光周期有關 ,它是感受延長光照信號所必需的。而 PhyB 與短日植物(SDP)誘導光周期有關。最近 ,王偉等在光敏核不育水稻中的研究說明 , PhyA 在暗期中的顯著增加可能是誘導育性轉變的光周期反應所必需的。
近些年來 ,隨著分子生物學的日益興起和發展 ,人們也開始從基因分子水平來闡明光敏色素的作用機制。許多實驗都證明 ,光敏色素對基因表達的調控大都在轉錄水平上進行的。人們通過許多實驗證明 ,從光敏色素的活化到基因表達之間存在著一系列的信號傳導中間體。而且 ,人們也利用生物化學的手段證明 ,G蛋白、cGMP、 cAMP 磷脂酶、 IP3、 Ca2 +等都是光敏色素信號傳遞鏈的組分。Molchan[25 ]對燕麥幼苗的研究 ,紅光和遠紅光都影響了燕麥的 cAMP 水平。Trctyn[26 ]提出 ,依賴 Ca2 +的信號傳導途徑可能是 Phy 作用機制的一個典型例子。而Anccta也指出 ,Phy 和鈣調蛋白的磷酸化有關。最近 ,Quail的研究也表明 ,不同 Phy 的功能可能是由將信號途徑特定地分向不同家族成員所導致的。Sharma研究認為 ,Phy作為一種絲氨酸激酶而使其自身磷酸化或使其它蛋白磷酸化而調節基因的轉錄水平。Quail利用酵母雜交實驗 ,證明 Phy 經光轉變后 ,可以從溶質轉到核內 ,在核內與螺旋2環2螺旋蛋白結合 ,直接與特定基因的啟動子發生作用 ,而 Ferenc等也確實證實 ,光質和光強能控制光敏色素的核輸入。近年來的一個重大進展是 Ni . M等發現的蛋白因子 PIF3(轉錄調節因子) ,它與 PhyB 的 Pf r 直接結合 ,控制基因的表達 ,而且 ,他還發現 ,PhyB N 末端的生色團結合區和C末端二聚體區對 PIF3 的結合力很弱 ,好像與結合PIF3 的長度有關。Neuhaus曾利用功能互補實驗 ,分離到一個 17bp 的天冬醯氨合成酶基因(ASI)的順式作用啟動子 ,而這種啟動子是 Phy 調節 ASI 所必需的 ,並且 ,它的活性是受 Ca2 +和cGMP所調節的。最近 ,Frankhauser在擬南芥中發現了一種作用因子 RSF1 ,它被認為是幼苗 PhyA介導的光信號傳導的一個組分 ,它對 PhyA 的穩定性和生物合成沒有影響 ,但是 ,在成熟的植物中 ,RSF1 的缺乏並不影響擬南芥的光周期開花反應。Deng X. W等發現了光敏色素作用因子 COP1 ,它是由三種特殊的結構域組成 ,並含有核定位信號(NLS)和細胞質定位信號(CLS) ,在光下 ,COP1 可以從核內轉到細胞質 ,在暗中 ,它卻存在於核內抑制光形態建成。同時 ,Stacey的實驗進一步證實 ,COP1 在亞細胞內的定位變化是 NLS 和 CLS 協調作用的結果。Hsieh的報道也指出 ,COP1 是生長素調節反應和光敏色素信號傳導的共同組分 ,PhyA 介導的遠紅光對 COP1有抑制作用 ,但是卻受生長素的促進。Colon2Carmon2a同樣也證明 ,Phy對生長素( IAA)蛋白的磷酸化可能是 Phy和 IAA 調控植物發育的分子機制。
美國馬里蘭州貝爾茨維爾(Beltsville)農業研究中心的Borthwick和Hendricks(1952)以大型光譜儀將白光分離成單色光,處理萵苣種子,發現紅光(波長650~680nm)促進種子發芽,而遠紅光(波長710~740nm)逆轉這個過程。1959年Butler等研製出雙波長分光光度計,測定黃化玉米幼苗的吸收光譜。他們發現,經紅光處理后,幼苗的吸收光譜中的紅光區域減少,而遠紅光區域增多;如果用遠紅光處理,則紅光區域增多,遠紅光區域消失。紅光和遠紅光輪流照射后,這種吸收光譜可多次的可逆變化。上述結果說明這種紅光-遠紅光可逆反應的受體可能是具兩種形式的單一色素。他們以後成功地分離出這種吸收紅光-遠紅光可逆轉換的光受體(色素蛋白質),稱之為光敏色素(phytochrome)。
光敏色素在植物體內至少存在兩種類型(Furuya,1993):一種在黃化幼苗中含量較高,在黑暗中才能合成,而在光下不穩定,成為黃化組織光敏色素(etiolate tissue phytochome,Phy Ⅰ),它的吸收峰在666nm;另一種以綠色組織為主,在光下相對穩定,且在光下和暗中均可合成,稱為綠色組織光敏色素(green tissue phytochome,PhyⅡ),吸收峰652nm。分子生物學實驗表明,被子植物中存在光敏色素基因(稱作PHY)家族。例如玉米的PHY數目2~4個,燕麥的PHY數超過4個。在擬南芥幼苗中發現了5種不同的光敏色素基因,分別被命名為PHYA、PHYB、PHYC、PHYD、PHYE。其中PHYA編碼的蛋白phyA屬PhyⅠ型光敏色素,接收波長700~750nm連續遠紅光,對光不穩定,在光下,其mRNA的活性受到抑制。其餘四種基因編碼的蛋白phyB、phyC、phyD、phyE屬光敏PhyⅡ型光敏色素,具有高度的光穩定性,不受光的影響,接收600~700nm紅光,屬組成型表達。至今已證明,PHYA、PHYB基因編碼的蛋白可組裝成全光敏色素phyA、phyB,phyA主要控制遠紅光對幼苗下胚軸的伸長作用,而phyB主要控制紅光對幼苗下胚軸的抑制作用。
存在於高等植物的所有部分,是植物體本身合成的一種調節生長發育的色蛋白。由蛋白質及生色團2部分組成,後者是4個吡咯分子連接成直鏈,與藻膽素類似。所有具光合作用的植物(光合細菌除外)均含有,含量極低。從不同植物中分離出的光敏色素,分子量範圍為120~127千道爾頓。有2種類型(近代研究認為還有若干中間型):一為紅光吸收型(Pr),最大吸收峰在666納米;另一為遠紅光吸收型(Pfr),最大吸收峰在730納米,兩者可以很快地相互轉變,Pr是生理失活型。它以Pr狀態合成,並在黑暗中積累,所以黃化幼苗中有Pr無Pfr。在紅光或白光照射下,大多數Pr轉變為Pfr。Pfr可發生降解、在暗中緩慢地逆轉為Pr及參與反應。因此,Pr在光中的總量比暗中少得多,有實驗指出,在暗中生長的雙子葉植物和單子葉植物幼苗中的含量比生長在光下的幼苗高出30至100倍,在純溶液中,用紅光照射后,Pfr為81%,Pr為19%。遠紅光照射后,幾乎所有Pfr轉變為Pr,Pfr僅留存2%。在日光下由於Pr吸收紅光比Pfr吸收遠紅光更為有效,所以在自然光照下,Pfr比Pr為多,Pfr約佔總量的60%。2者的比例隨太陽的入射角、雲層厚薄及地上林冠的變化而變化。曾有人指出生理活躍型的Pfr與另一未知物(X)形成複合物[Pfr·X]。由於X化學性質以及*2者比例的不同,[Pfr·X]複合物將引起種種生理反應,如控制開花、打破某些需光種子休眠、下胚軸彎鉤的伸長、幼葉和子葉變綠及展開、含羞草的感震運動和抑制花色素苷的形成等。 Pr 紅光→ Pfr 遠紅光→ Pr。
光敏色素的生理作用甚為廣泛,它影響植物一生的形態建成,從種子萌發到開花、結果及衰老。
高等植物中一些由光敏色素控制的反應。
1.種子萌發 6.小葉運動 11.光周期 16.葉脫落。
2.彎鉤張開 7.滲透性 12.花誘導 17.塊莖形成。
3.節間延長 8.向光敏感性 13.子葉張開 18.性別表現。
4.根原基起始 9.花色素形成 14.肉質化 19.葉片張開(單)。
5.分化和擴大 10.質體形成 15.偏上性 20.節律現象。
光敏色素接受光刺激到發生反應的時間有快有慢。快反應以秒計,如棚田效應(Tanada effect)和轉板藻葉綠體運動。棚田效應指離體綠豆根尖在紅光下誘導膜產生少量正電荷,所以能黏附在帶負電的玻璃表面,而遠紅光則逆轉這種黏附現象。慢反應則以小時和天數計,例如,紅光促進萵苣種子萌發和誘導幼苗黃化反應。
基本信息
- 中文名
- 光敏色素
- 外文名
- phytochrome
- 本質
- 光受體(色素蛋白質)
- 分佈
- 植物各個器官中
- 細胞中
- 胞質溶膠和細胞核中都有
- 功能
- 吸收紅光-遠紅光可逆轉換