核酸
許多核苷酸聚合成的生物大分子化合物
核酸是由許多核苷酸聚合成的生物大分子化合物,為生命的最基本物質之一。核酸廣泛存在於所有動植物細胞、微生物體內,生物體內的核酸常與蛋白質結合形成核蛋白。不同的核酸,其化學組成、核苷酸排列順序等不同。根據化學組成不同,核酸可分為核糖核酸(簡稱RNA)和脫氧核糖核酸(簡稱DNA)。DNA是儲存、複製和傳遞遺傳信息的主要物質基礎。RNA在蛋白質合成過程中起著重要作用——其中轉運核糖核酸,簡稱tRNA,起著攜帶和轉移活化氨基酸的作用;信使核糖核酸,簡稱mRNA,是合成蛋白質的模板;核糖體的核糖核酸,簡稱rRNA,是細胞合成蛋白質的主要場所。核酸由核苷酸組成,而核苷酸單體由五碳糖、磷酸基和含氮鹼基組成。如果五碳糖是核糖,則形成的聚合物是RNA;如果五碳糖是脫氧核糖,則形成的聚合物是DNA。
高分子化合物的一類,是生命的最基本物質之一。由許多(至少幾十)個核苷酸通過磷酸二酯鍵連接而成,存在於所有動植物細胞、微生物和病毒、噬菌體內,對生物的生長、遺傳、變異等現象起著決定作用。根據所含成分,可分為核糖核酸(RNA)和脫氧核糖核酸(DNA)兩類。核酸與遺傳、腫瘤的發生、病毒的感染性、射線對機體的作用都有重要關係。
核酸同蛋白質一樣,也是生物大分子。核酸的相對分子質量很大,一般是幾十萬至幾百萬。核酸水解后得到許多核苷酸,實驗證明,核苷酸是組成核酸的基本單位,即組成核酸分子的單體。一個核苷酸分子是由一分子含氮的鹼基、一分子五碳糖和一分子磷酸組成的。根據五碳糖的不同可以將核苷酸分為脫氧核糖核苷酸和核糖核苷酸。
核酸大分子可分為兩類:脫氧核糖核酸(DNA)和核糖核酸(RNA),在蛋白質的複製和合成中起著儲存和傳遞遺傳信息的作用。核酸不僅是基本的遺傳物質,而且在蛋白質的生物合成上也占重要位置,因而在生長、遺傳、變異等一系列重大生命現象中起決定性的作用。
| 核酸 | DNA | RNA |
|---|---|---|
| 名稱 | 脫氧核糖核酸 | 核糖核酸 |
| 結構 | 規則的雙螺旋結構 | 通常呈單鏈結構 |
| 基本單位 | 脫氧核糖核苷酸 | 核糖核苷酸 |
| 五碳糖 | 脫氧核糖 | 核糖 |
| 含氮鹼基 | A(腺嘌呤) G(鳥嘌呤) C(胞嘧啶) T(胸腺嘧啶) | A(腺嘌呤) G(鳥嘌呤) C(胞嘧啶) U(尿嘧啶) |
| 分佈 | 主要存在於細胞核,少量存在於線粒體和葉綠體 | 主要存在於細胞質 |
| 功能 | 攜帶遺傳信息,在生物體的遺傳、變異和蛋白質的生物合成中具有極其重要的作用 | 作為遺傳物質:只在RNA病毒中;不作為遺傳物質:在DNA控制蛋白質合成過程中起作用。mRNA是蛋白質是合成的直接模板、tRNA能攜帶特定氨基酸、rRNA是核糖體的組成成分;催化作用:酶的一種 |
(1)五碳糖——DNA是脫氧核糖;RNA是核糖。
(2)鹼基——DNA是A、T、C、G(不含U);RNA是A、U、C、G(不含T)。
(3)DNA通常是雙螺旋結構;RNA通常是單鏈,局部可形成雙螺旋結構。
核酸在實踐應用方面有極重要的作用,現已發現近2000種遺傳性疾病都和DNA結構有關。如人類鐮刀形紅血細胞貧血症是由於患者的血紅蛋白分子中一個氨基酸的遺傳密碼發生了改變,白化病患者則是DNA分子上缺乏產生促黑色素生成的酪氨酸酶的基因所致。腫瘤的發生、病毒的感染、射線對機體的作用等都與核酸有關。70年代以來興起的遺傳工程,使人們可用人工方法改組DNA,從而有可能創造出新型的生物品種。如應用遺傳工程方法已能使大腸桿菌產生胰島素、干擾素等珍貴的生化藥物。
1869年,F.Miescher從膿細胞中提取到一種富含磷元素的酸性化合物,因存在於細胞核中而將它命名為“核質”(nuclein)。但核酸(nucleic acids)這一名詞在Miescher發現“核質”20年後才被正式啟用,當時已能提取不含蛋白質的核酸製品。早期的研究僅將核酸看成是細胞中的一般化學成分,沒有人注意到它在生物體內有什麼功能這樣的重要問題。
1944年,Avery等為了尋找導致細菌轉化的原因,他們發現從S 型肺炎球菌中提取的DNA與R型肺炎球菌混合后,能使某些R型菌轉化為S型菌,且轉化率與DNA純度呈正相關,若將DNA預先用DNA酶降解,轉化就不發生。結論是:S型菌的DNA將其遺傳特性傳給了R型菌,DNA就是遺傳物質。從此核酸是遺傳物質的重要地位才被確立,人們把對遺傳物質的注意力從蛋白質移到了核酸上。
核酸研究中劃時代的工作是Watson和Crick於1953年創立的DNA 雙螺旋結構模型。模型的提出建立在對DNA下列三方面認識的基礎上:
1.核酸化學研究中所獲得的DNA化學組成及結構單元的知識,特別是Chargaff於1950~1953年發現的DNA化學組成的新事實;DNA中四種鹼基的比例關係為A/T=G/C=1。
2.X線衍射技術對DNA結晶的研究中所獲得的一些原子結構的最新參數。
3.遺傳學研究所積累的有關遺傳信息的生物學屬性的知識。綜合這三方面的知識所創立的DNA雙螺旋結構模型,不僅闡明了DNA分子的結構特徵,而且提出了DNA作為執行生物遺傳功能的分子,從親代到子代的DNA複製 (replication)過程中,遺傳信息的傳遞方式及高度保真性。其正確性於1958年被Meselson和Stahl的著名實驗所證實。DNA雙螺旋結構模型的確立為遺傳學進入分子水平奠定了基礎,是現代分子生物學的里程碑。從此核酸研究受到了前所未有的重視。
DNA分子具有規則的雙螺旋結構。是由兩條相互平行且反向右旋的脫氧核苷酸長鏈所構成,分子中央的鹼基鹼基互補配對原則以氫鍵相連。DNA獨特的雙螺旋結構和鹼基互補配對能力使DNA的兩條鏈“可分”,“可合”,半保留複製自如,“精確”複製的DNA通過細胞分裂等方式傳遞下去,使子代(或體細胞)含有與親代相似的遺傳物質。但“精確”複製並不是絕對不存在差錯,複製差錯率非常低(約1~10億分之一),然而卻導致基因發生突變,出現新基因,產生可遺傳的變異,有利於生物的進化。
瑞士生物學家:米舍爾Friedrich Miescher
美國生物學家:沃森Watson,James Dewey
英國生物物理學家:克里克Crick,Francis Harry Compton
英國醫生:格里菲思Griffith, F.
阿委瑞Avery O. T.
赫爾希Hershey, A.D.
蔡斯Chase, M.
30多年來,核酸研究的進展日新月異,所積累的知識幾年就要更新。其影響面之大,幾乎涉及生命科學的各個領域,現代分子生物學的發展使人類對生命本質的認識進入了一個嶄新的天地。
核酸
遺傳信息以核苷酸順序的形式貯存在DNA分子中,它們以功能單位在染色體上佔據一定的位置構成基因(gene)。因此,搞清DNA順序無疑是非常重要的。1975年 Sanger發明的DNA測序(DNAsequencing)加減法為實現這一企圖起了關鍵性的作用。由此而發展起來的大片段DNA順序快速測定技術──Maxam 和Gilbert的化學降解法(1977年)和Sanger的末端終止法(1977年),已是核酸結構與功能研究中不可缺少的分析手段。
中國學者洪國藩於1982年提出了非隨機的有序DNA測序新策略,對DNA測序技術的發展作出了重要貢獻。憑藉先進的DNA測序技術及其它基因分析手段,人類正在進行一項以探明自身基因組(genome)全部核苷酸順序(單倍基因組 含3×10鹼基對)為目標的宏偉計劃──人類基因組圖譜製作計劃(human genome mapping project)。據稱,此項計劃的實現,將對全人類的健康產生無止境的影響。
Watson-Crick模型創立36年後的1989年,一項新技術──掃描隧道顯微鏡(scanning tummeling microscopy,STM)使人類首次能直接觀測到近似自然環境中的單個DNA分子的結構細節,觀測數據的計算 機處理圖像能在原子級水平上精確度量出DNA分子的構型、旋轉周期、大溝(major groove)及小溝(minor groove)。這一成果是對DNA 雙螺旋結構模型真實性的最直接而可信的證明。此項技術無疑會對人類最終完全解開遺傳之謎提供有力的幫助。可喜的是,中國科學家在這項世界領先的研究中也佔有一席之地。
核酸分子通常很大。實際上,DNA分子可能是已知的最大的單個生物分子。
但也有比較小的核酸分子。
核酸分子的大小範圍從21個核苷酸(小干擾RNA)到大染色體(人類染色體是一個含有2.47億個鹼基對的單個分子)不等。
核酸完全水解產生嘌呤和嘧啶等鹼性物質、戊糖(核糖或脫氧核糖)和磷酸的混合物。核酸部分水解則產生核酸和核苷酸。每個核苷分子含一分子鹼基和一分子戊糖,一分子核苷酸部分水解后除產生核苷外,還有一分子磷酸。
核酸中的糖和磷酸鹽通過磷酸二酯鍵以交替鏈(糖 - 磷酸骨架)相互連接。磷酸基團所連接的碳是糖的3'-末端,與碳原子結合的碳是5'-末端,這就產生了核酸的方向性。核鹼基通過N-糖苷鍵與糖連接。
在RNA和DNA中也發現了非標準核苷,它們通常來自DNA分子內的標準核苷或初始RNA轉錄物的修飾。轉移RNA(tRNA)分子含有特別多的修飾核苷。
天然存在的DNA分子在大多數情況下是雙鏈的,而RNA分子是單鏈的。然而,有許多例外。一些病毒具有由雙鏈RNA構成的基因組,而其他病毒具有單鏈DNA基因組,並且在某些情況下,可形成具有三個或四個鏈的核酸結構。
核酸是生物體內的高分子化合物。它包括脫氧核糖核酸(deoxyribonucleicacid,DNA)和核糖核酸(ribonucleicacid,RNA)兩大類。
單個核苷酸是由含氮有機鹼(稱鹼基)、戊糖(即五碳糖)和磷酸三部分構成的。
鹼基(base):構成核苷酸的鹼基分為嘌呤(purine)和嘧啶>;(pyrimi-dine)二類。前者主要指腺嘌呤(adenine,A)和鳥嘌呤(guanine,G),DNA和RNA中均含有這二種鹼基。後者主要指胞嘧啶(cytosine,C)胸腺嘧啶(thymine,T)和尿嘧啶(uracil,U),胞嘧啶存在於DNA和RNA中,胸腺嘧啶只存在於DNA中,尿嘧啶則只存在於RNA中。
嘌呤環上的N-9或嘧啶環上的N-1是構成核苷酸時與核糖(或脫氧核糖)形成糖苷鍵的位置。
此外,核酸分子中還發現數十種修飾鹼基(themodifiedcomponent),又稱稀有鹼基,(unusualcomponent)。它是指上述五種鹼基環上的某一位置被一些化學基團(如甲基化、甲硫基化等)修飾后的衍生物。一般這些鹼基在核酸中的含量稀少,在各種類型核酸中的分佈也不均一。如DNA中的修飾鹼基主要見於噬菌體DNA,RNA中以tRNA含修飾鹼基最多。
戊糖(五碳糖):RNA中的戊糖是D-核糖(即在2號位上連接的是一個羥基),DNA中的戊糖是D-2-脫氧核糖(即在2號位上只連一個H)。D-核糖的C-2所連的羥基脫去氧就是D-2脫氧核糖。
戊糖C-1所連的羥基是與鹼基形成糖苷鍵的基團,糖苷鍵的連接都是β-構型。
核苷(nucleoside):由D-核糖或D-2脫氧核糖與嘌呤或嘧啶通過糖苷鍵連接組成的化合物。核酸中的主要核苷有八種。
核苷酸(nucleotide):核苷酸與磷酸殘基構成的化合物,即核苷的磷酸酯。核苷酸是核酸分子的結構單元。核酸分子中的磷酸酯鍵是在戊糖C-3’和C-5’所連的羥基上形成的,故構成核酸的核苷酸可視為3’-核苷酸或5’-核苷酸。DNA分子中是含有A,G,C,T四種鹼基的脫氧核苷酸;RNA分子中則是含A,G,C,U四種鹼基的核苷酸。
當然核酸分子中的核苷酸都以形式存在,但在細胞內有多種遊離的核苷酸,其中包括一磷酸核苷、二磷核苷和三磷酸核苷。
3’,5’-磷酸二酯鍵:核酸是由眾多核苷酸聚合而成的多聚核苷酸(polynucleotide),相鄰二個核苷酸之間的連接鍵即:3’,5’-磷酸二酯鍵。這種連接可理解為核苷酸糖基上的3'位羥基與相鄰5'核苷酸的磷酸殘基之間,以及核苷酸糖基上的5'位羥基與相鄰3'核苷酸的磷酸殘基之間形成的兩個酯鍵。多個核苷酸殘基以這種方式連接而成的鏈式分子就是核酸。無論是DNA還是RNA,其基本結構都是如此,故又稱DNA鏈或RNA鏈。DNA鏈的結構如下示意圖。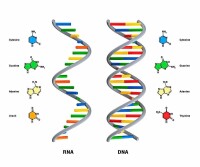
核酸
寡核苷酸(oligonucleotide):這是與核酸有關的文獻中經常出現的一個術語,一般是指二至十個核苷酸殘基以磷酸二酯鍵連接而成的線性多核苷酸片段。但在使用這一術語時,對核苷酸殘基的數目並無嚴格規定,在不少文獻中,把含有三十甚至更多個核苷酸殘基的多核苷酸分子也稱作寡核苷酸。寡核苷酸可由儀器自動合成,它可作為DNA合成的引物(primer)、基因探針(probe)等,在現代分子生物學研究中具有廣泛的用途。
核酸鏈的簡寫式:核酸分子的簡寫式是為了更簡單明了的敘述高度複雜的核酸分子而使用的一些簡單表示式。它所要表示的主要內容是核酸鏈中的核苷酸(或鹼基)。下面介紹二種常用的簡寫式。
字元式:書寫一條多核苷酸鏈時,用英文大寫字母縮寫符號代表鹼基(DNA和RNA中所含主要鹼基及縮寫符號見表1-1),用小寫英文字母P代表磷酸殘基。核酸分子中的糖基、糖苷鍵和酯鍵等均省略不寫,將鹼基和磷酸相間排列即可。因省略了糖基,故不再註解“脫氧”與否,凡簡寫式中出現T就視為DNA鏈,出現U則視為RNA鏈。以5'和3'表示鏈的末端及方向,分別置於簡寫式的左右二端。下面是分別代表DNA鏈和RNA鏈片段的二個簡寫式:
5'pApCpTpTpGpApApCpG3'DNA
5'pApCpUpUpGpApApCpG3'RNA
此式可進一步簡化為:
5'pACTTGAACG3'
5'pACUUGAACG3'
上述簡寫式的5'-末端均含有一個磷酸殘基(與糖基的C-5'位上的羥基相連),3'-末端含有一個自由羥基(與糖基的C-3'位相連),若5'端不寫P,則表示5'-末端為自由羥基。雙鏈DNA分子的簡寫式多採用省略了磷酸殘基的寫法,在上述簡式的基礎上再增加一條互補鏈(complentarystrand)即可,鏈間的配對鹼基用短縱線相連或省略,錯配(mismatch)鹼基對錯行書寫在互補鏈的上下兩邊,如下所示:
5'GGAATCTCAT3'
3'CCTTAGAGTA5'
5'GGAATC錯配)
線條式:在字元書寫基礎上,以垂線(位於鹼基之下)和斜線(位於垂線與P之間)分別表示糖基和磷酸酯鍵。
上式中,斜線與垂線部的交點為糖基的C-3'位,斜線與垂線下端的交點為糖基的C-5'位。這一書寫式也可用於表示短鏈片段。不難看出,簡寫式表示的中心含義就是核酸分子的一級結構,即核酸分子中的核苷酸(或鹼基)排列順序。
從前面的描述我們也可以看得很清楚,核酸氧化分解后變成了磷酸和鹼基的嘌呤和嘧啶,嘌呤也是導致人類尿酸增高和痛風的主要原因。
核酸氧化分解—生成嘌呤—嘌呤在肝臟進一步氧化成為(2,6,8—三氧嘌呤)又稱為尿酸,尿酸鹽沉積到關節腔等組織引起痛風發作。
因此,核酸不是越多越好,同時,這也說明了為什麼中老年易患痛風,因為年紀大了,大量的細胞死亡,而細胞內有大量的核酸,生成嘌呤,再生成尿酸,從而導致痛風發作。防治好痛風就是要防止核酸被氧化。
核酸(nucleic acid)是重要的生物大分子,它的構件分子是核苷酸(nucleotide)。
天然存在的核酸可分為:
⑴脫氧核糖核酸(deoxyribonucleic acid,DNA)和核糖核酸(ribonucleic acid,RNA)
DNA貯存細胞所有的遺傳信息,是物種保持進化和世代繁衍的物質基礎。
RNA中參與蛋白質合成的有三類:
轉移RNA(transfer RNA,tRNA)、核糖體RNA(ribosomal RNA,rRNA)和信使RNA(messenger RNA,mRNA)
20世紀末,發現許多新的具有特殊功能的RNA,幾乎涉及細胞功能的各個方面。
核苷酸可分為:核糖核苷酸(RNA的構件分子)和 脫氧核糖核苷酸(DNA構件分子)
細胞內還有各種遊離的核苷酸和核苷酸衍生物,它們具有重要的生理功能。
核苷酸由 核苷(nucleoside)磷酸(Phosphonic.acid)組成
核苷由:鹼基(base)和 戊糖(Pentose)組成
構成核苷酸中的鹼基是含氮雜環化合物,由嘧啶(pyrimidine)和嘌呤(purine)構成。
核酸:1.嘌呤鹼:腺嘌呤(A)鳥嘌呤(G)2.嘧啶鹼:胞嘧啶(C)胸腺嘧啶(T) 尿嘧啶(U)
DNA中含有4種鹼基:腺嘌呤、鳥嘌呤和胞嘧啶,胸腺嘧啶主要存在於DNA中。
RNA中含也有4種鹼基:腺嘌呤、鳥嘌呤和胞嘧啶,尿嘧啶主要存在於RNA中。
在某些tRNA分子中也有胸腺嘧啶,少數幾種噬菌體的DNA含尿嘧啶而不是胸腺嘧啶。這五種鹼基受介質pH的影響出現酮式、烯醇式互變異構體。
在DNA和RNA中,尤其是tRNA中還有一些含量甚少的鹼基,稱為稀有鹼基(rare bases)稀有鹼基種類很多,大多數是甲基化鹼基。tRNA中含稀有鹼基高達10%。
核酸中有兩種戊糖DNA中為D-2-脫氧核糖(D-2-deoxyribose),RNA中則為D-核糖(D-ribose)。在核苷酸中,為了與鹼基中的碳原子編號相區別核糖或脫氧核糖中碳原子標以C-1’,C-2’等。脫氧核糖與核糖兩者的差別只在於脫氧核糖中與2’位碳原子連結的不是羥基而是氫,這一差別使DNA在化學上比RNA穩定得多。
核苷是戊糖與鹼基之間以糖苷鍵(glycosidic bond)相連接而成。戊糖中C-1’與嘧啶鹼的N-1或者與嘌吟鹼的N9相連接,戊糖與鹼基間的連接鍵是N-C鍵,一般稱為N-糖苷鍵。
RNA中含有稀有鹼基,並且還存在異構化的核苷。如在tRNA和rRNA中含有少量假尿嘧啶核苷(用ψ表示),在它的結構中戊糖的C-1不是與尿嘧啶的N-1相連接,而是與尿嘧啶C-5相連接。
核苷中的戊糖5’碳原子上羥基被磷酸酯化形成核苷酸。核苷酸分為核糖核苷酸與脫氧核糖核苷酸兩大類。依磷酸基團的多少,有一磷酸核苷、二磷酸核苷、三磷酸核苷。核苷酸在體內除構成核酸外,尚有一些遊離核苷酸參與物質代謝、能量代謝與代謝調節,如三磷酸腺苷(ATP)是體內重要能量載體;三磷酸尿苷參與糖原的合成;三磷酸胞苷參與磷脂的合成;環腺苷酸(cAMP)和環鳥苷酸(cGMP)作為第二信使,在信號傳遞過程中起重要作用;核苷酸還參與某些生物活性物質的組成:如尼克醯胺腺嘌呤二核苷酸(NAD+),尼克醯胺腺嘌呤二核苷酸磷酸(NADP+)和黃素腺嘌呤二核苷酸(FAD)。
核酸是由核苷酸聚合而成的生物大分子。組成DNA的脫氧核糖核苷酸主要是dAMP、dGMP、dCMP和dTMP,組成RNA的核糖核苷酸主要是AMP、GMP、CMP和UMP。核酸中的核苷酸以3’,5’磷酸二酯鍵構成無分支結構的線性分子。核酸鏈具有方向性,有兩個末端分別是5’末端與3’末端。5’末端含磷酸基團,3’末端含羥基。核酸鏈內的前一個核苷酸的3’羥基和下一個核苷酸的5’磷酸形成3’,5’磷酸二酯鍵,故核酸中的核苷酸被稱為核苷酸殘基。通常將小於50個核苷酸殘基組成的核酸稱為寡核苷酸(oligonucleotide),大於50個核苷酸殘基稱為多核苷酸(polynucleotide)。
(一)DNA的二級結構
DNA二級結構即雙螺旋結構(double helix structure)。20世紀50年代初Chargaff等人分析多種生物DNA的鹼基組成發現的規則。
DNA雙螺旋模型的提出不僅揭示了遺傳信息穩定傳遞中DNA半保留複製的機制,而且是分子生物學發展的里程碑。
DNA雙螺旋結構特點如下:①兩條DNA互補鏈反向平行。②由脫氧核糖和磷酸間隔相連而成的親水骨架在螺旋分子的外側,而疏水的鹼基對則在螺旋分子內部,鹼基平面與螺旋軸垂直,螺旋旋轉一周正好為10個鹼基對,螺距為3.4nm,這樣相鄰鹼基平面間隔為0.34nm並有一個36°的夾角。③DNA雙螺旋的表面存在一個大溝(major groove)和一個小溝(minor groove),蛋白質分子通過這兩個溝與鹼基相識別。④兩條DNA鏈依靠彼此鹼基之間形成的氫鍵而結合在一起。根據鹼基結構特徵,只能形成嘌呤與嘧啶配對,即A與T相配對,形成2個氫鍵;G與C相配對,形成3個氫鍵。因此G與C之間的連接較為穩定。⑤DNA雙螺旋結構比較穩定。維持這種穩定性主要靠鹼基對之間的氫鍵以及鹼基的堆集力(stacking force)。
生理條件下,DNA雙螺旋大多以B型形式存在。右手雙螺旋DNA除B型外還有A型、C型、D型、E型。此外還發現左手雙螺旋Z型DNA。Z型DNA是1979年Rich等在研究人工合成的CGCGCG的晶體結構時發現的。Z-DNA的特點是兩條反向平行的多核苷酸互補鏈組成的螺旋呈鋸齒形,其表面只有一條深溝,每旋轉一周是12個鹼基對。研究表明在生物體內的DNA分子中確實存在Z-DNA區域,其功能可能與基因表達的調控有關。DNA二級結構還存在三股螺旋DNA,三股螺旋DNA中通常是一條同型寡核苷酸與寡嘧啶核苷酸-寡嘌呤核苷酸雙螺旋的大溝結合,三股螺旋中的第三股可以來自分子間,也可以來自分子內。三股螺旋DNA存在於基因調控區和其他重要區域,因此具有重要生理意義。
(二)DNA三級結構——超螺旋結構
DNA三級結構是指DNA鏈進一步扭曲盤旋形成超螺旋結構。生物體內有些DNA是以雙鏈環狀DNA形式存在,如有些病毒DNA,某些噬菌體DNA,細菌染色體與細菌中質粒DNA,真核細胞中的線粒體DNA、葉綠體DNA都是環狀的。環狀DNA分子可以是共價閉合環,即環上沒有缺口,也可以是缺口環,環上有一個或多個缺口。在DNA雙螺旋結構基礎上,共價閉合環DNA(covalently close circular DNA)可以進一步扭曲形成超螺旋形(super helical form)。根據螺旋的方向可分為正超螺旋和負超螺旋。正超螺旋使雙螺旋結構更緊密,雙螺旋圈數增加,而負超螺旋可以減少雙螺旋的圈數。幾乎所有天然DNA中都存在負超螺旋結構。
(三)DNA的四級結構——DNA與蛋白質形成複合物
在真核生物中其基因組DNA要比原核生物大得多,如原核生物大腸桿菌的DNA約為4.7×10kb,而人的基因組DNA約為3×10 kb,因此真核生物基因組DNA通常與蛋白質結合,經過多層次反覆摺疊,壓縮近10 000倍后,以染色體形式存在於平均直徑為5μm的細胞核中。線性雙螺旋DNA摺疊的第一層次是形成核小體(nucleosome)。猶如一串念珠,核小體由直徑為11nm×5.5nm的組蛋白核心和盤繞在核心上的DNA構成。核心由組蛋白H2A、H2B、H3和H4各2分子組成,為八聚體,146 bp長的DNA以左手螺旋盤繞在組蛋白的核心1.75圈,形成核小體的核心顆粒,各核心顆粒間有一個連接區,約有60 bp雙螺旋DNA和1個分子組蛋白H1構成。平均每個核小體重複單位約佔DNA 200 bp。DNA組裝成核小體其長度約縮短7倍。在此基礎上核小體又進一步盤繞摺疊,最後形成染色體。
(四)DNA結構的多態性
Watson和Crick所推導出來的DNA結構在生物學研究中有深遠意義。他們是以在生理鹽溶液中抽出的DNA纖維在92%相對溫度下進行X-射線衍射圖譜為依據進行推設的。在這一條件下得出的DNA稱B構象。實際上在溶液中的DNA的確呈這一構象,這也是最常見的DNA構象。但是,研究表明DNA的結構是動態的。在以鈉、鉀或銫作反離子,相對溫度為75%時,DNA分子的X-射線衍射圖給出的是A構象。這一構象不僅出現於脫水DNA中,還出現在RNA分子中的雙螺旋區域的DNA-RNA雜交分子中。如果以鋰作反離子,相對溫度進一步降為66%,則DNA是C構象。但是這一構象僅在實驗室中觀察到,還未在生物體中發現。這些DNA分子中G-C鹼基對較少,這些分子將取D和E構象。這些研究表明DNA的分子結構不是一成不變的,在不同的條件下可以有所不同。但是,這些不同構象的DNA都有共同的一點,即它們都是右手雙螺旋;兩條反向平行的核苷酸鏈通過Watson-Crick鹼基配對結合在一起;鏈的重複單位是單核苷酸;這些螺旋中都有兩個螺旋溝,分為大溝與小溝,只是它們的寬窄和深淺程度有所不同。
但是,Wang和Rich等人在研究人工合成的CGCGCG單晶的X-射線衍射圖譜時分別發現這種六聚體的構象與上面講到的完全不同。它是左手雙螺旋,在主鏈中各個磷酸根呈鋸齒狀排列,有如“之”字形一樣,因此叫它Z構象(英文字Zigzag的第一個字母)。還有,這一構象中的重複單位是二核苷酸而不是單核苷酸;而且Z-DNA只有一個螺旋溝,它相當於B構象中的小溝,它狹而深,大溝則不復存在。
立即就有幾個問題被提了出來:這種結構是怎樣生成的?這一結構在天然狀態下存在嗎?它有什麼生物學意義?
研究表明,Z-DNA的形成是DNA單鏈上出現嘌呤與嘧啶交替排列所成的。比如CGCGCGCG或者CACACACA。這種鹼基排列方式會造成核苷酸的糖苷鍵的順式和反式構象的交替存在。當鹼基與糖構成反式結構時,它們之間離得遠;而當它們成順式時,就彼此接近。嘧啶糖苷鍵通常是反式的,而嘌呤糖苷酸鍵既可成順式的也可成反式的。而在Z-DNA中,嘌呤鹼是順式的。這樣,在Z-DNA中嘧啶的糖苷鏈離開小溝向外挑出,而嘌呤上的糖苷鍵則彎向小溝。嘌呤與嘧啶的交替排列就使得糖苷鍵也是順式與反式交替排列,從而使Z-DNA主鏈呈鋸齒狀或“之”字形。
人們相信,並用實驗證明細胞DNA分子中確實存在有Z-DNA區。而且,細胞內有一些因素可以促使B-DNA轉變為Z-DNA。比如,胞嘧啶第五位碳原子的甲基化,在甲基周圍形成局部的疏水區。這一區域擴伸到B-DNA的大溝中,使B-DNA不穩定而轉變為Z-DNA。這種C5甲基化現象在真核生物中是常見的。因此在生物B構象的DNA中某些區段具有Z-DNA構象是可能的。DNA真是一個構象可變動態分子。
Z-DNA有會么生物學意義呢?應當指出Z-DNA的形成通常在熱力學上是不利的。因為Z-DNA中帶負電荷的磷酸根距離太近了,這會產物靜電排斥。但是,DNA鏈的局部不穩定區的存在就成為潛在的解鏈位點。DNA解螺旋卻是DNA複製和轉錄等過程中必要的環節,因此認為這一結構位點與基因調節有關。比如SV40增強子區中就有這種結構,又如鼠類微小病毒DNS複製區起始點附近有GC交替排列序列。此外,DNA螺旋上溝的特徵在其信息表達過程中起關鍵作用。調控蛋白都是通過其分子上特定的氨基酸側鏈與DNA雙螺旋溝中的鹼基對一側的氫原子供體或受體相互作用,形成氫鍵從而識別DNA上的遺傳信息的。大溝所帶的遺傳信息比小溝多。溝的寬窄和深淺也直接影響到調控蛋白質對DNA信息的識別。Z-DNA中大溝消失,小溝狹而深,使調探蛋白識別方式也發生變化。這些都暗示Z-DNA的存在不僅僅是由於DNA中出現嘌呤-啶嘧交替排列之結果,也一定是在漫漫的進化長河中對DNA序列與結構不斷調整與篩選的結果,有其內在而深刻的含意,只是人們還未充分認識而已。
DNA構象的可變性,或者說DNA二級結構的多態性的發現拓寬了人們的視野。原來,生物體中最為穩定的遺傳物質也可以採用不同的姿態來實現其豐富多彩的生物的奧妙,也讓人們在這一領域中探索和攀越時減少疲勞和厭倦,樂而忘返,從而有更多更新的發現。
多年來,DNA結構的研究手段主要是X射線衍線技術,其結果是通過間接觀測多個DNA分子有關結構參數的平均值而獲得的。同時,這項技術的樣品分析條件使被測DNA分子與天然狀態相差甚遠。因此,在反映DNA結構真實性方面這種方法存在著缺陷。1989年,應用掃描隧道顯微鏡(STM)研究DNA結構克服了上述技術的缺陷。這種先進的顯微技術,不僅可將被測物放大500萬倍,且能直接觀測接近天然條件下單個DNA分子的結構細節。應該說它所取得的DNA結構資料更具有"權威性"。表1-6是STM測到的B-DNA結構參數及其與X射線衍線資料的比較結果。STM研究還證實了d(CG)重複序列的寡核苷酸片段為Z-DNA結構的事實。STM技術的應用是DNA結構研究中的重要進展,可望在探索DNA結構的某些未知點上展示巨大潛力。
(一)基因(gene)的現代分子生物學概念是指能編碼有功能的蛋白質多肽鏈或合成RNA所必需的全部核酸序列,是核酸分子的功能單位。一個基因通常包括編碼蛋白質多肽鏈或RNA的編碼序列,保證轉錄和加工所必需的調控序列和5’端、3’端非編碼序列。另外在真核生物基因中還有內含子等核酸序列。
(二)基因組(genome)是指一個細胞或病毒所有基因及間隔序列,儲存了一個物種所有的遺傳信息。在病毒中通常是一個核酸分子的鹼基序列,單細胞原核生物是它僅有的一條染色體的鹼基序列,而多細胞真核生物是一個單倍體細胞內所有的染色體。如人單倍體細胞的23條染色體的鹼基序列。多細胞真核生物起源於同一個受精卵,其每個體細胞的基因組都是相同的。
● ● 病毒基因組
● ● 原核生物基因組
● ● 真核生物基因組
在高等真核生物中基因序列占整個基因組不到10%,大部分是非編碼的間隔序列。人類基因組研究結果發現在人的基因組中與蛋白質合成有關的基因只佔整個基因組2 %。真核生物基因組的最大的特點是出現分隔開的基因,在這類基因中有編碼作用的序列稱外顯子(exon),沒有編碼作用的序列稱內含子(intron),它們彼此間隔排列。
絕大部分RNA分子都是線狀單鏈,但是RNA分子的某些區域可自身回折進行鹼基互補配對,形成局部雙螺旋。在RNA局部雙螺旋中A與U配對、G與C配對,除此以外,還存在非標準配對,如G與U配對。RNA分子中的雙螺旋與A型DNA雙螺旋相似,而非互補區則膨脹形成凸出(bulge)或者環(loop),這種短的雙螺旋區域和環稱為髮夾結構(hairpin)。髮夾結構是RNA中最普通的二級結構形式,二級結構進一步摺疊形成三級結構,RNA只有在具有三級結構時才能成為有活性的分子。RNA也能與蛋白質形成核蛋白複合物,RNA的四級結構是RNA與蛋白質的相互作用。
(一) tRNA的結構
tRNA約佔總RNA的15%,tRNA主要的生理功能是在蛋白質生物合成中轉運氨基酸和識別密碼子,細胞內每種氨基酸都有其相應的一種或幾種tRNA,因此tRNA的種類很多,在細菌中約有30~40種tRNA,在動物和植物中約有50~100種tRNA。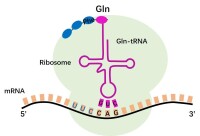 1. tRNA一級結構:
1. tRNA一級結構:
核酸
tRNA是單鏈分子,含73~93核苷酸,分子質量為24 000~31 000kDa,沉降係數4S。含有10%的稀有鹼基。如二氫尿嘧啶(DHU)、核糖胸腺嘧啶(rT)和假尿苷(ψ)以及不少鹼基被甲基化,其3’端為CCA-OH,5’端多為pG,分子中大約30%的鹼基是不變的或半不變的,也就是說它們的鹼基類型是保守的。
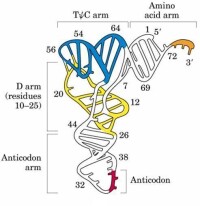
2. tRNA二級結構: tRNA二級結構為三葉草型(如右圖)。配對鹼基形成局部雙螺旋而構成臂,不配對的單鏈部分則形成環。三葉草型結構由4臂4環組成。氨基酸臂由7對鹼基組成,雙螺旋區的3’末端為一個4個鹼基的單鏈區-NCCA-OH 3’,腺苷酸殘基的羥基可與氨基酸α羧基結合而攜帶氨基酸。二氫尿嘧啶環以含有2個稀有鹼基二氫尿嘧啶(DHU)而得名,不同tRNA其大小並不恆定,在8~14個鹼基之間變動,二氫尿嘧啶臂一般由3~4對鹼基組成。反密碼環由7個鹼基組成,大小相對恆定,其中3個核苷酸組成反密碼子(anticodon),在蛋白質生物合成時,可與mRNA上相應的密碼子配對。反密碼臂由5對鹼基組成。額外環在不同tRNA分子中變化較大可在4~21個鹼基之間變動,又稱為可變環,其大小往往是tRNA分類的重要指標。TψC環含有7個鹼基,大小相對恆定,幾乎所有的tRNA在此環中都含TψC序列,TψC臂由5對鹼基組成。
3. tRNA的三級結構:
核酸
(二)mRNA
原核生物中mRNA轉錄后一般不需加工,直接進行蛋白質翻譯。mRNA轉錄和翻譯不僅發生在同一細胞空間,而且這兩個過程幾乎是同時進行的。真核細胞成熟mRNA是由其前體核內不均一RNA(heterogeneous nuclear RNA,hnRNA)剪接並經修飾后才能進入細胞質中參與蛋白質合成。所以真核細胞mRNA的合成和表達發生在不同的空間和時間。mRNA的結構在原核生物中和真核生物中差別很大。下面分別作一介紹:
1. 原核生物mRNA結構特點
原核生物的mRNA結構簡單,往往含有幾個功能上相關的蛋白質的編碼序列,可翻譯出幾種蛋白質,為多順反子。在原核生物mRNA中編碼序列之間有間隔序列,可能與核糖體的識別和結合有關。在5’端與3’端有與翻譯起始和終止有關的非編碼序列,原核生物mRNA中沒有修飾鹼基,5’端沒有帽子結構,3’端沒有多聚腺苷酸的尾巴(polyadenylate tail,polyA尾巴)。原核生物的mRNA的半衰期比真核生物的要短得多,轉錄后1min,mRNA降解就開始。
2. 真核生物mRNA結構特點
真核生物mRNA為單順反子結構,即一個mRNA分子只包含一條多肽鏈的信息。在真核生物成熟的mRNA中5’端有m7GpppN的帽子結構,帽子結構可保護mRNA不被核酸外切酶水解,並且能與帽結合蛋白結合識別核糖體並與之結合,與翻譯起始有關。3’端有polyA尾巴,其長度為20~250個腺苷酸,其功能可能與mRNA的穩定性有關,少數成熟mRNA沒有polyA尾巴,如組蛋白mRNA,它們的半衰期通常較短。
(三)rRNA的結構
rRNA占細胞總RNA的80%左右,rRNA分子為單鏈,局部有雙螺旋區域具有複雜的空間結構,原核生物主要的rRNA有三種,即5S、16S和23S rRNA,如大腸桿菌的這三種rRNA分別由120、1542和2904個核苷酸組成。真核生物則有4種,即5S、5.8S、18S和28S rRNA,如小鼠,它們相應含121、158、1874和4718個核苷酸。rRNA分子作為骨架與多種核糖體蛋白(ribosomal protein)裝配成核糖體。
所有生物體的核糖體都由大小不同的兩個亞基所組成。原核生物核糖體為70S,由50S和30S兩個大小亞基組成。30S小亞基含16S的rRNA和21種蛋白質,50S大亞基含23S和5S兩種rRNA及34種蛋白質。真核生物核糖體為80S,是由60S和40S兩個大小亞基組成。40S的小亞基含18S rRNA及33種蛋白質,60S大亞基則由28S、5.8S和5S 3種rRNA及49種蛋白質組成。
(四)其他RNA分子
20世紀80年代以後由於新技術不斷產生,人們發現RNA有許多新的功能和新的RNA基因。細胞核內小分子RNA(small nuclear RNA,snRNA)是細胞核內核蛋白顆粒(Small nuclear ribonucleoprotein particles,snRNPs)的組成成分,參與mRNA前體的剪接以及成熟的mRNA由核內向胞漿中轉運的過程。核仁小分子RNA(small nucleolar RNA,snoRNA)是類新的核酸調控分子,參與rRNA前體的加工以及核糖體亞基的裝配。胞質小分子RNA(small cytosol RNA, scRNA)的種類很多,其中7S LRNA與蛋白質一起組成信號識別顆粒(signal recognition particle,SRP),SRP參與分泌性蛋白質的合成,反義RNA(antisense RNA)由於它們可以與特異的mRNA序列互補配對,阻斷mRNA翻譯,能調節基因表達。核酶是具有催化活性的RNA分子或RNA片段。在醫學研究中已設計了針對病毒的致病基因mRNA的核酶,抑制其蛋白質的生物合成,為基因治療開闢新的途徑,核酶的發現也推動了生物起源的研究。微RNA(microRNA,miRNA)是一種具有莖環結構的非編碼RNA,長度一般為20-24個核苷酸,在mRNA翻譯過程中起到開關作用,它可以與靶mRNA結合,產生轉錄后基因沉默作用(post-transcriptional gene silencing,PTGS),在一定條件下能釋放,這樣mRNA又能翻譯蛋白質,由於miRNA的表達具有階段特異性和組織特異性,它們在基因表達調控和控制個體發育中起重要作用。
隨著基因組研究不斷深入,蛋白組學研究逐漸展開,RNA的研究也取得了突破性的進展,發現了許多新的RNA分子,人們逐漸認識到DNA是攜帶遺傳信息分子,蛋白質是執行生物學功能分子,而RNA即是信息分子,又是功能分子。人類基因組研究結果表明,在人類基因組中約有30000~40000個基因,其中與蛋白質生物合成有關的基因只佔整個基因組的2%,對不編碼蛋白質的98%基因組的功能有待進一步研究,為此20世紀末科學家在提出蛋白質組學后,又提出RNA組學。RNA組是研究細胞的全部RNA基因和RNA的分子結構與功能。
①酸效應:在強酸和高溫,核酸完全水解為鹼基,核糖或脫氧核糖和磷酸。在濃度略稀的的無機酸中,最易水解的化學鍵被選擇性的斷裂,一般為連接嘌呤和核糖的糖苷鍵,從而產生脫嘌呤核酸。
②鹼效應
1. DNA:當PH值超出生理範圍(pH7~8)時,對DNA結構將產生更為微妙的影響。鹼效應使鹼基的互變異構態發生變化。這種變化影響到特定鹼基間的氫鍵作用,結果導致DNA雙鏈的解離,稱為DNA的變性
2.RNA:PH較高時,同樣的變性發生在RNA的螺旋區域中,但通常被RNA的鹼性水解所掩蓋。這是因為RNA存在的2`-OH參與到對磷酸脂鍵中磷酸分子的分子內攻擊,從而導致RNA的斷裂。
③化學變性:一些化學物質能夠使DNA/RNA在中性PH下變性。由堆積的疏水鹼基形成的核酸二級結構在能量上的穩定性被削弱,則核酸變性。
①黏性:DNA的高軸比等性質使得其水溶液具有高黏性,很長的DNA分子又易於被機械力或超聲波損傷,同時黏度下降。
② 浮力密度:可根據DNA的密度對其進行純化和分析。在高濃度分子質量的鹽溶液(CsCl)中,DNA具有與溶液大致相同的密度,將溶液高速離心,則CsCl趨於沉降於底部,從而建立密度梯度,而DNA最終沉降於其浮力密度相應的位置,形成狹帶,這種技術成為平衡密度梯度離心或等密度梯度離心。
③穩定性:核酸的結構相當穩定,其主要原因有1、鹼基對間的氫鍵2、鹼基的堆積作用3、環境中的陽離子。
①減色性:dsDNA相對於ssDNA是減色的,而ssDNA相對於dsDNA是增色的。
② DNA純度:A260/A280。
①熱變性:dsDNA與RNA的熱力學表現不同,隨著溫度的升高RNA中雙鏈部分的鹼基堆積會逐漸地減少,其吸光性值也逐漸地,不規則地增大。較短的鹼基配對區域具有更高的熱力學活性,因而與較長的區域相比變性快。而dsDNA熱變性是一個協同過程。分子末端以及內部更為活躍的富含A-T的區域的變性將會使其赴京的螺旋變得不穩定,從而導致整個分子結構在解鏈溫度下共同變性。
② 復性:DNA的熱變性可通過冷卻溶液的方法復原。不同核酸鏈之間的互補部分的復性稱為雜交。
一般來說,進化程度高的生物DNA分子應越大,能貯存更多遺傳信息。但進化的複雜程度與DNA大小並不完全一致,如哺乳類動物DNA約為3×10 bp,但有些兩棲類動物、南美肺魚DNA大小可達1010bp到1011bp。
常用測定DNA分子大小的方法有電泳法、離心法。凝膠電泳是當前研究核酸的最常用方法,凝膠電泳有瓊脂糖(agarose)凝膠電泳和聚丙烯醯胺(polyacrylamide)凝膠電泳。
DNA和RNA中的糖苷鍵與磷酸酯鍵都能用化學法和酶法水解。在很低pH條件下DNA和RNA都會發生磷酸二酯鍵水解。並且鹼基和核糖之間的糖苷鍵更易被水解,其中嘌呤鹼的糖苷鍵比嘧啶鹼的糖苷鍵對酸更不穩定。在高pH時,RNA的磷酸酯鍵易被水解,而DNA的磷酸酯鍵不易被水解。
水解核酸的酶有很多種,若按底物專一性分類,作用於RNA的稱為核糖核酸酶(ribonuclease,RNase),作用於DNA的則稱為脫氧核糖核酸酶(deoxyribonuclease,DNase)。按對底物作用方式分類,可分核酸內切酶(endonuclease)與核酸外切酶(exonuclease)。核酸內切酶的作用是在多核苷酸內部的3’,5’磷酸二酯鍵,有些內切酶能識別DNA雙鏈上特異序列並水解有關的3’,5’磷酸二酯鍵。核酸內切酶是非常重要的工具酶,在基因工程中有廣泛用途。而核酸外切酶只對核酸末端的3’,5’磷酸二酯鍵有作用,將核苷酸一個一個切下,可分為5’→3’外切酶,與3’→5’外切酶。
在一定理化因素作用下,核酸雙螺旋等空間結構中鹼基之間的氫鍵斷裂,變成單鏈的現象稱為變性(denaturation)。引起核酸變性的常見理化因素有加熱、酸、鹼、尿素和甲醯胺等。在變性過程中,核酸的空間構象被破壞,理化性質發生改變。由於雙螺旋分子內部的鹼基暴露,其A260值會大大增加。A260值的增加與解鏈程度有一定比例關係,這種關係稱為增色效應(hyperchromic effect)。如果緩慢加熱DNA溶液,並在不同溫度測定其A260值,可得到“S”形DNA熔化曲線(melting curve)。從DNA熔化曲線可見DNA變性作用是在一個相當窄的溫度內完成的。
當A260值開始上升前DNA是雙螺旋結構,在上升區域分子中的部分鹼基對開始斷裂,其數值隨溫度的升高而增加,在上部平坦的初始部分尚有少量鹼基對使兩條鏈還結合在一起,這種狀態一直維持到臨界溫度,此時DNA分子最後一個鹼基對斷開,兩條互補鏈徹底分離。通常把加熱變性時DNA溶液A260升高達到最大值一半時的溫度稱為該DNA的熔解溫度(melting temperature Tm),Tm是研究核酸變性很有用的參數。Tm一般在85~95℃之間,Tm值與DNA分子中G C含量成正比。
變性DNA在適當條件下,可使兩條分開的單鏈重新形成雙螺旋DNA的過程稱為復性(renaturation)。當熱變性的DNA經緩慢冷卻后復性稱為退火(annealing)。DNA復性是非常複雜的過程,影響DNA復性速度的因素很多:DNA濃度高,復性快;DNA分子大復性慢;高溫會使DNA變性,而溫度過低可使誤配對不能分離等等。最佳的復性溫度為Tm減去25℃,一般在60℃左右。離子強度一般在0.4mol/L以上。
具有互補序列的不同來源的單鏈核酸分子,按鹼基配對原則結合在一起稱為雜交(hybridization)。雜交可發生在DNA-DNA、RNA-RNA和DNA-RNA之間。雜交是分子生物學研究中常用的技術之一,利用它可以分析基因組織的結構,定位和基因表達等,常用的雜交方法有Southern印跡法,Northern印跡法和原位雜交(insitu hybridization)等。
核酸、蛋白質誰更“牛”?
一般人都知道,生命是蛋白質存在的形式,蛋白質是生命的基礎。在發現核酸前,這句話是對的,但當核酸被發現后,應該說最本質的生命物質是核酸,或是把上述的這句話更正為核酸是生命的基礎。按照現代生物學的觀點,蛋白體是包括核酸和蛋白質的生物大分子。
核酸在生命中為什麼比蛋白質更重要呢?因為生命的重要性是能自我複製,而核酸就能夠自我複製。蛋白質的複製是根據核酸所發出的指令,使氨基酸根據其指定的種類進行合成,然後再按指定的順序排列成所需要複製的蛋白質。世界上各種有生命的物質都含有蛋白體,蛋 白體中有核酸和蛋白質,還沒有發現有蛋白質而沒有核酸的生命。但在有生命的病毒研究中,卻發現病毒以核酸為主體,蛋白質和脂肪以及脂蛋白等只不過充作其外殼,作為與外界環境的界限而已,當它鑽入寄生細胞繁殖子代時,把外殼留在細胞外,只有核酸進入細胞內,並使細胞在核酸控制下為其合成子代的病毒。這種現象,美國科學家比喻為人和汽車的關係。即把核酸比為人,蛋白質比作汽車,人駕駛汽車到處跑,外表上看,人車一體是有生命運動的東西,而真正的生命是人,汽車只是由人製造的載入的外殼。有一種類病毒,是能繁殖子代的有生命物體,其中只有核酸而沒蛋白質,可見核酸是真正的生命物質。
因此中國1996年最新出版的《人體生理學》改變了舊教科書中只提蛋白質是生命基礎的缺陷,明確提出:“蛋白質和核酸是一切生命活動的物質基礎。”
然而,多少年來,人們在一味追求蛋白質、維生素、微量元素等營養時,卻把最重要的角色——核酸忘卻了,這不能不說是人類生命史上的一大遺憾。
沒有核酸,就沒有蛋白,也就沒有生命。
核酸也稱多聚核苷酸,是由許多個核苷酸聚合而成的生物大分子,核苷酸是由含氮的鹼基、核糖或脫氧核糖、磷酸三種分子連接而成。鹼基與糖通過糖苷鍵連接成核苷,核苷與磷酸以酯鍵連接成核苷酸。核苷酸是生物體內一類重要含氮化合物,是各種核酸的基本組成單位。根據核酸所含戊糖的不同,可分為核糖核酸(RNA)和脫氧核糖核酸(DNA)二種。
核酸不但是一切生物細胞的基本成分,還對生物體的生長、發育、繁殖、遺傳及變異等重大生命現象起主宰作用。它在生物科學的地位,可用“沒有核酸就沒有生命”這句話來概括。
飲食核酸的營養保健作用如下:
1.飲食核酸與免疫 ‘
從核酸對機體各系統的影響來看,免疫系統是最敏感也是最直接受影響的系統。1985年科學家就證實無核酸飲食或低核酸飲食配方飼餵的實驗動物,其細胞免疫功能低下,條件致病菌就可使其感染。無核酸飲食致使T淋巴細胞發育障礙、功能低下,而沒有細胞免疫反應的發生,同時影響T細胞依賴的體液免疫的產生;補充核酸營養后可恢復免疫系統的發育和免疫功能。實驗表明,核酸是維持機體正常免疫功能和免疫系統生長代謝的必需營養物質。
2.飲食核酸與衰老和內分泌
衰老是機體各組織器官的退行性變化,關於衰老發生機制的學說很多,如自由基學說、免疫學說、內分泌學說、遺傳學說等。脂質過氧化隨年齡增大而增高,並伴有酶與非酶系統防禦功能下降,導致體內自由基濃度升高。代謝性、退行性疾病的發生和發展與體內過氧化脂質含量高度正相關。飲食核酸作為使遺傳物質活潑代謝的原料,具有極強的抗生物氧化、消除體內自由基和全面增強免疫功能及性激素分泌的作用,因此在延緩衰老方面優勢顯著。
3.飲食核酸與增殖細胞
飲食中補加核酸有助於肝臟再生和受損傷的小腸恢復功能。有無核酸飲食對比研究證明,一段時期內膳食中如缺乏核酸,將對大鼠肝臟的超微結構及功能造成不良影響,提示飲食核酸是維持肝臟處於正常生理狀態的必需營養物質。血液中的紅細胞、白細胞、血小板和血漿蛋白等也都是代謝較快的增殖細胞系,加之它們中的大多數均無從頭合成核酸的能力,因此它們的代謝和功能也都需要充足的核酸營養。再生障礙性貧血和抗癌藥物、放療、化療等引起的貧血,即缺鐵性貧血之外的貧血均需補充核酸營養,以改善骨髓造血功能和血液成分的代謝活力。
4.飲食核酸與癌症
人體每日約有數百萬個癌狀細胞出現,它們幾乎全部被機體的免疫監視系統和核酸、維生素等食物成分,在形成大的癌細胞克隆前排除掉。因此在日常生活中盡量避免致癌因子的作用,增加核酸等防癌因素的作用非常必要。
5.飲食核酸與痴獃等神經障礙
食物核酸提取物對痴呆症狀的改善非常令人鼓舞。在大鼠實驗中,如給大鼠腦注射RNA合成阻斷劑,則所掌握的學習能力和記憶能力在5小時后喪失,但如在注射RNA合成阻斷劑的同時注射拮抗阻斷劑的物質,這種記憶喪失就不發生。美國哈佛大學的研究也表明,老年痴獃患者腦內神經纖維變化多的部位,RNA和蛋白質合成顯著減少,因此發生記憶障礙。
6.飲食核酸與循環系統
核酸營養對循環系統的作用是抑制過氧化脂質的形成,抑制膽固醇的生成,擴張血管,改善血流,糾正心肌代償不良,促進血管壁再生,抑制血小板凝集i因此核酸被認為對腦血栓、心肌梗死、高血壓和動脈粥樣硬化症有較好的營養保健作用。
7、飲食核酸與糖尿病
非胰島素依賴性糖尿病與生活方式和運動不足關係密切,如果在普通的飲食療法的基礎上,再加上核酸飲食,將收到更好的效果。其原因:一是糖尿病患者血清中過氧化脂質增多,核酸及其代謝產物對其具有較強的清除作用;二是由於核酸的促細胞(包括促胰髒的胰島素分泌細胞)代謝功能。除此之外,核酸的代謝產物腺苷還有抑製糖的分解作用,使糖在小腸內的吸收減緩。
除上述作用外,飲食核酸還有以下作用:減肥,提高機體對環境變化的耐受力,顯著的抗疲勞、增強機體對冷熱的抵抗力、促進攝人氧氣的利用,促進小鼠生殖系統的發育等。
對於嬰兒、迅速成長期的孩子、老年體弱多病、全身感染、外傷手術者、肝功能不全以及白細胞、T細胞、淋巴細胞降低人群等,可以額外補充核酸類物質。世界衛生組織規定,每天膳食中核酸的量不大於2克,扣除食物中的核酸攝入量,每天補充小於1.5克核酸是合適的。
人工核酸治療白血病
日本工業技術院產業技術融合領域研究所已開發出了治療白血病的人造核酸。科研小組的成員、東京大學研究生院教授多比良和誠根據動物實驗結果認為,這種人造核酸將來有望成為治療白血病的主要藥物。
這次研究的對象是慢性骨髓性白血病(MCL),患者的異常遺傳因子是由兩個正常的遺傳因子連接而成的,新開發的人造核酸可以發現這種變異遺傳基因並將其切斷。科學家過去也發現過能找到特定的遺傳因子序列並將其切斷的分子,但在切斷特定遺傳因子序列的同時往往對正常細胞造成傷害。而新開發出的核酸只在發現異常遺傳因子時才被激活,平時則潛伏不動。
科研小組用人體白血病細胞進行了動物實驗。他們將可與人造核酸反應的細胞和不可與人造核酸反應的細胞分別注射到8隻實驗鼠的體內。移植后第13周時,不與人造核酸反應的細胞全部死亡,而與人造核酸反應的細胞全部存活,證明人造核酸在生物體內十分有效。
科研小組說,此人造核酸的臨床應用尚有諸多問題要解決,將來很可能是把患者的骨髓細胞抽出來,經人造核酸處理后,再把正常細胞的骨髓輸回患者體內。
核酸的生物功能
核酸在生物體內主要與蛋白質合成核蛋白存在,它既是蛋白質生物合成不可缺少的物質,又是生物遺傳的物質基礎。
DNA主要存在於細胞核中,它們是遺傳信息的攜帶者,DNA的結構決定生物合成蛋白質的特定結構,並保證這種遺傳特性傳給下一代。RNA主要存在於細胞質中,它們是以DNA為模板而形成的,並且直接參加蛋白質的生物合成過程。因此,DNA是RNA的模板,而RNA又是蛋白質的模板。存在於DNA分子上的遺傳信息就是這樣由DNA傳遞給RNA,再傳遞給蛋白質。通過DNA 複製,遺傳信息一代代傳下去,正因為有這樣的功能,人們把核酸譽為“生命之源”和“生命之本”。
基本信息
- 中文名
- 核酸
- 外文名
- nucleic acid
- 拼音
- hé suān
- 相對分子質量
- 一般是幾十萬至幾百萬
- 分類
- 核糖核酸、脫氧核糖核酸
- EINECS
- 277-256-77
- CAS號
- 63231-63-0