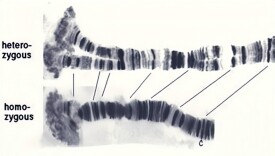
多線染色體
多線染色體
多線染色體(polytene chromosome)一種纜狀的巨大染色體,見於有些生物生命周期的某些階段里的某些細胞中。由核內有絲分裂產生的多股染色單體平行排列而成。
是指在某些細胞中,由於核內DNA複製多次而細胞不分裂,產生的子染色體并行排列,加之體細胞同源染色單體配對並緊密結合在一起,阻止了染色質進一步包裝,從而形成由許多條相同染色質纖維并行排列成體積很大的多線染色體。
各染色單體上的染色粒(見燈刷染色體)並排排列,構成多線染色體的帶,帶與帶之間則稱間帶。多線染色體的這種結構可用光學顯微鏡觀察,也能在多線染色體上用原位分子雜交法進行基因定位,並就其結構與功能之間的關係進行系統研究,因此是細胞學和遺傳學研究的有用材料。
核內DNA多次複製產生的子染色體平行排列,且體細胞內同源染色體配對,緊密結合在一起,從而阻止了染色體纖維進一步聚縮,形成體積很大的由多條染色體組成的結構叫多線染色體。多線化的細胞處於永久間期,體積也相應增大,它存在於雙翅目昆蟲的幼蟲組織內,如唾液腺、氣管等。
多線染色體不是生長到一定程度就進入有絲分裂,而是不斷生長,繼續複製,而且新的複製體總是沿其全長整齊地與原來的染色體並列著的,因而染色體就生長得極其龐大。例如,在果蠅唾腺細胞中每一個多線染色體都是經過大約9個循環的複製產生的,所以每條多線染色體至少包含了500-1000條單染色體(DNA纖絲),某些昆蟲的多線染色體包含了多達16000條。經過醋酸洋紅或地衣紅染色后,在高倍光鏡下就可以看到每條多線染色體都是由暗帶和明間帶直線交替組成的。同時也已證明,大部分DNA存在於暗區帶之內,每條區帶都相應於染色體上染色粒的聚合區域,它能被鹼性染料染得很深,孚爾根染色呈現陽性,而明間帶則幾乎不著色。以後又證明了每條區帶都包括幾個或幾十個基因位點。
1881年E.G.巴爾比安尼首先在雙翅目搖蚊(Chironomus)幼蟲的唾腺細胞中觀察到多線染色體,但未引起注意。1933年在遺傳學成就的影響下T.S.佩因特在果蠅唾腺,E.海茨和H.鮑爾等在毛蚊屬(Bibio)再次看到這種染色體后,人們才予重視。此後在昆蟲的多種組織如腸、氣管、脂肪體細胞和馬爾皮基氏管上皮細胞內以及在其他動植物的一些高度特化細胞如某些原生動物及附子屬(Aconitum)植物的反足細胞里也發現了這種巨大染色體。
用顯微鏡觀察多線染色體,只須把腺體等材料放在一滴固定劑和染色劑的混合液中用玻片壓破腺體細胞核,即可看到多線染色體分散出來。
多線染色體上有按一定次序排列的橫帶,一個帶含有多個基因。帶的數目,大小,位置隨不同的多線染色體而不同,因此可以據此鑒別染色體。多線染色體的帶型在光鏡下可見。
多線染色體上還可見膨脹而成的小泡,即疏鬆區。疏鬆區是緊密纏繞的DNA分子鬆開,即基因正在活動的部分,故疏鬆區總能檢測到RNA。
并行排列的染色質纖維
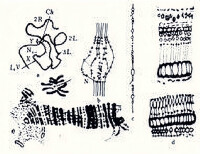
圖1
同種動物的不同組織以及不同動物的相同組織的多線化程度各不相同。例如搖蚊馬爾皮基氏管細胞的染色體最多能複製 9次,從而使每對同源染色體組含有 512(29)條染色單體;唾腺細胞染色體複製13次,因而含有8192(213)條染色單體。
帶和間帶
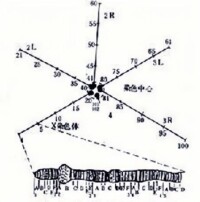
圖2
各種多線染色體上帶的數目、形態、大小及其分佈位置都很穩定。帶和間帶的分布圖譜簡稱帶譜。C.B.布里奇斯首先建立起黑腹果蠅唾腺染色體的帶譜。至今已知該帶的總數約有5000多條。圖2示4條染色體,分別命名為X;2L(左臂)和2R(右臂);3L和3R;4。從X染色體的遠端開始,依次將全部染色體分成102個區域。每一個區域可再進一步劃分成A、B、C等小區域(圖2)。從左邊開始可將每個小區域里的帶、間帶和脹泡依次編號。不同帶的形態和大小變化很大,同一條染色體在不同情況下其帶譜不同。其他一些物種的詳細帶譜也有報道。
帶譜與遺傳圖譜
把用雜交試驗得到的果蠅多線染色體的遺傳圖譜與正常的帶譜比較,可以看出每條帶相當於一個遺傳單位,並且可以鑒定出許多具有特殊遺傳功能的帶的位置。在特殊情況下,一條帶可能同時有幾個結構基因。例如,用原位雜交法曾經證明5SRNA基因的大部分拷貝位於2R的一條帶(56F)上;組蛋白mRNA只能雜交到2L的兩條帶和它們之間的間帶(39D-39E)上。
脹泡與基因表達
在個體發育的某個階段或某些化學物質的誘發下,多線染色體的某些帶紋變得疏鬆膨大而形成脹泡。最大的脹泡叫做巴爾比安尼氏環。脹泡是基因轉錄和翻譯的形態學標誌,在這裡DNA解旋呈開放環,RNA的合成很活躍;核糖體排列成多聚核糖體長鏈,多肽鏈的長度有一個梯度,甚至還可觀察到從巴爾比安尼環上新合成的蛋白質分泌顆粒。
在一種搖蚊 (C.pallidivittatus)唾腺前葉細胞的染色體4上,近著絲點處有巴爾比安尼環4,這些細胞能分泌特異的蛋白質顆粒。然而,在另一種搖蚊(C.tentanus)中則沒有巴爾比安尼環4,也沒有這種分泌顆粒。這兩種動物的雜交試驗表明,編碼分泌顆粒蛋白質的結構基因位於染色體4的巴爾比安尼環4上。
在尖眼蕈蚊 (Sciara coprophila)等動物還發現有一種非常特殊的脹泡──DNA脹泡。3H-胸腺嘧啶核苷和3H-尿嘧啶核苷能摻入這種脹泡。它們是額外的DNA複製區域(圖3),其轉錄活性和5-甲基胞嘧啶含量之間有反相關性。
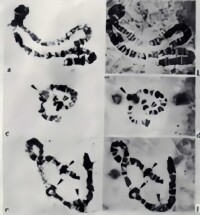
多線染色體
如果在蛋白質合成受到抑制的條件下(例如用放線菌酮等),使唾腺受到激素處理,仍能誘發早期脹泡,但不能誘發晚期脹泡。這說明早期脹泡的形成不需要蛋白質合成,晚期脹泡的形成可能是早期脹泡基因產物作用的後果。早期脹泡的活性始終依賴於蛻皮激素,一旦除去激素后脹泡就萎縮;而晚期脹泡在沒有激素存在時仍能正常地出現,也說明這一情況。此外,早期和晚期脹泡對蛻皮激素濃度的反應不同,早期脹泡74EF起反應的範圍很寬(可相差500倍,但晚期脹泡(62E)起反應的範圍較窄(僅相差4倍)。
果蠅幼蟲唾腺經高溫(例如37℃,40分鐘)處理后多線染色體出現明顯的變化,許多脹泡萎縮,原來活躍的基因被抑制。與此同時,一組特異基因被激活,從而出現一些特異性脹泡。這一現象叫做熱休克,所產生的脹泡叫做熱休克脹泡(或稱熱誘發脹泡)。脹泡的大小與熱休克處理的激烈程度有關。
果蠅多線染色體對熱休克刺激的反應發生在基因表達的不同水平上。在轉錄(即RNA合成)水平,至少有3種反應:①誘發合成一組熱休克RNA,其中有些能被翻譯成熱休克蛋白質;②抑制了在25℃時進行的大多數其他mRNA的合成;③中斷了5S和18S+28S核糖體RNA基因轉錄產物的正常加工。
用3H尿苷摻入多線染色體的放射自顯影試驗,以及抗 RNA聚合酶Ⅱ抗體和抗染色體蛋白質的單克隆抗體在多線染色體上的分佈的定位研究表明,在一般情況下,脹泡和間帶是 RNA合成的活躍場所。然而經熱休克處理后,RNA 聚合酶Ⅱ在轉錄上活躍的熱休克脹泡處迅速積累起來,對染色體蛋白質專一的單克隆抗體的染色體上的分佈也集中在熱休克脹泡上,同時從其他區域上消失,這說明熱休克脹泡是轉錄和轉譯的活躍場所。熱休克處理后在轉移水平上所發生的變化,一方面是誘發少數熱休克多肽的合成,另一方面是抑制了原來在25℃培養時出現的大多數蛋白質的合成。熱休克開始后10分鐘,用35S-甲硫氨酸摻入法和SDS-聚丙烯醯胺凝膠電泳並將它們分成幾條帶。有關熱休克多肽的確切數目,比較一致的看法是,在黑腹果蠅中主要有8種不同的多肽,而在果蠅中有6種多肽。現已有直接證據表明,特異的熱休克脹泡能編碼特殊的熱休克蛋白質。
由於多線染色體上的脹泡的變化直接反映了基因轉錄活性的變化,因此隨著克隆重組DNA和原位分子雜交技術的發展,對多線染色體的研究必將為揭示真核生物基因的調控機製作出貢獻。