共找到2條詞條名為DNA複製的結果 展開
- DNA雙鏈進行的複製過程
- 1997年出品美國電影
DNA複製
DNA雙鏈進行的複製過程
DNA複製是指DNA雙鏈在細胞分裂以前進行的複製過程,從一個原始DNA分子產生兩個相同DNA分子的生物學過程。DNA複製是通過名為半保留複製的機制來得以順利完成的。
DNA複製發生在所有DNA為遺傳物質的生物體中,是生物遺傳的基礎。
DNA複製
DNA複製主要包括引發、延伸、終止三個階段。
DNA複製是生物遺傳的基礎,是所有生物體中最基本的過程。而這一過程是半保留複製,是以最開始的雙鏈分子中的一條作為模板進行DNA複製,產生兩個完全一致的DNA分子。細胞水平的校正和糾錯機制能確保非常精確地複製DNA的拷貝。DNA複製發生在基因組的特定位置也就是起始點,DNA分子在起始點形成複製叉開始複製。
DNA複製從起始序列開始單向或雙向進行。合成DNA雙螺旋的兩條鏈是反向平行排列的,其中一條鏈的起始端與另一條鏈的末尾端平行排列在一起,每一個複製叉只有一條鏈是按照從尾到頭的正確方向指導新鏈從頭到尾方向合成。根據這條指導鏈,DNA複製持續向前合成複製叉。
DNA複製不能沿滯后鏈進行,也就是說,從頭到尾的DNA鏈,直到已經複製了足夠長度的DNA分子,否則DNA複製不會繼續沿著模本鏈進行複製,DNA複製於是從新合成複製叉處分開。在複製過程中必須暫停並等待更多的親本DNA鏈片段,而此時整個長度只是沿著開始到結束方向前進了一小段距離。
DNA複製為邊解旋邊複製,原核生物一般是單個複製起點,真核生物多個複製起點。
複製體是一個執行DNA複製的複雜分子機器。它由大量的次級元件組成,每一個次級元件在複製的過程中都行使一個特殊的功能。解螺旋酶能切斷兩條DNA分子之間的氫鍵,從而在DNA合成前分開兩條鏈。當解螺旋酶解開雙螺旋時,引導DNA其它區域的超螺旋體排列好。
旋轉酶的作用是解開由解旋酶切斷DNA鏈產生的超螺旋化,解旋酶使DNA鏈旋轉並釋放超螺旋體,使它們重新加入到DNA鏈中。旋轉酶最常見於複製叉的上游,形成超螺旋的位置。
由於DNA聚合酶只能連接DNA鏈(不能開始),所以由引物酶引導指導鏈進行複製。引物酶將與模本鏈互補的RNA引物加到DNA鏈上開始複製岡崎片段。
DNA合成酶Ⅲ由2個催化核心構成,一個引導DNA鏈複製,一個間隔DNA鏈。但是DNA合成酶Ⅲ不能停留足夠時間,有效地複製姐妹鏈。於是包含3個亞基的二聚物β聚合物共同包裹住DNA鏈使DNA合成酶Ⅲ留在DNA鏈上,確保DNA聚合酶Ⅲ能在鏈上合成幾千個核酸而不是幾百個。
DNA合成酶Ⅰ將引物酶添加的RNA引物去掉,完成岡崎片段。而DNA合成酶Ⅰ的作用會使岡崎片段之間產生小的空白區域,這就需要連接酶將岡崎片段連接起來,最終兩個岡崎片段的末端以共價鍵結合。
單鏈結合蛋白綁定在暴露的鹼基上竭力防止DNA鏈的不穩定並保證單鏈DNA之間不會由氫鍵形成危險的髮夾結構。DNA合成酶包含一個校對機制,通常指的是“外切核酸酶活性”,即將錯誤添加的核酸去除掉。
DNA聚合酶包含一個'校對'機制,通常被稱為 ‘外切酶活性'。這樣就刪除了誤添加的核苷酸。
DNA的複製是對那些堅持達爾文主義世界觀的的人們的一項基本挑戰。作為生物信息被複制並傳遞給後代的過程,這是一個對於細胞的自我複製過程必要的機制。細胞的自我複製對於任何選擇性的過程中都是必要的,比如自然選擇。因此,試圖用自然選擇來解釋這個機制巨大的複雜性需要人們先要假設他們想解釋的東西的客觀存在。由於其極為複雜的性質,大多數生化學者先前認為該系統產生,是在最後一個共同祖先的起源之前。此外,許多生化學家長久以來一直把在所有生命中觀察到的DNA複製的功能性的相似當作DNA複製的單一的起源。不過在1999年,美國國家衛生研究院的研究人員證明,參與細菌和古細菌或真核細胞(生命進化之樹的兩個主幹)的DNA複製的核心酶其實並沒有一個共同的進化起源。因此,它看起來好像細菌和古細菌獨自產生了兩個相同的DNA複製系統 — 在這兩個進化的譜系據信分化自最後的共同祖先之後。
認為這一工程奇迹是一次形成的就已令人驚嘆的,更不用說兩次。沒有明顯的原因表明DNA複製是通過一個半保留的,RNA引物依賴性的,雙向的機制發生的,該機制依靠前置鏈和滯后鏈產生DNA後代分子。即使DNA複製可以在兩個不同場合獨立地演變,考慮到他們的特性,有理由認為對於細菌和古細菌或真核細胞會出現根本不同的機制。但是沒有。
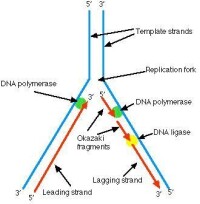
DNA複製
為什麼需要有RNA引物來引發DNA複製呢?這可能盡量減少DNA複製起始處的突變有關。DNA複製開始處的幾個核苷酸最容易出現差錯,因此,用RNA引物即使出現差錯最後也要被DNA聚合酶Ⅰ切除,提高了DNA複製的準確性。RNA引物形成后,由DNA聚合酶Ⅲ催化將第一個脫氧核苷酸按鹼基互補原則加在RNA引物3'-OH端而進入DNA鏈的延伸階段。
DNA雙螺旋的解旋
DNA在複製的時候,在DNA解旋酶的作用下,雙鏈首先解開,形成了複製叉,而複製叉的形成則是由多種蛋白質和酶參與的較複雜的複製過程
(1)單鏈DNA結合蛋白(single—stranded DNA binding protein,ssbDNA蛋白)
ssbDNA蛋白是較牢固結合在單鏈DNA上的蛋白質。原核生物ssbDNA蛋白和DNA結合時表現出協同效應:如果第一個ssbDNA蛋白結合到DNA上去能力為1,第二個的結合能力可高達103;真核生物細胞里的ssbDNA蛋白與單鏈DNA結合時則不表現上述效應。ssbDNA蛋白作用是保證解旋酶解開的單鏈在複製完成前能保持單鏈結構,以四聚體的形式存在於複製叉處,等待單鏈複製后才脫下來,重新循環。因此,ssbDNA蛋白僅保持單鏈的存在,是不起解旋作用。
(2)DNA解鏈酶(DNA helicase)
DNA解鏈酶可以通過水解ATP獲得能量以解開雙鏈DNA。這一種解鏈酶分解ATP的活性依賴於單鏈DNA的存在。若雙鏈DNA中有單鏈末端或切口,則DNA解鏈酶能首先結合在這一部分,然後逐步向雙鏈的方向移動。複製時,大部分DNA解旋酶沿滯后模板的5’—〉3’方向並隨著複製叉的前進而移動,只有個別解旋酶(Rep蛋白)是沿著3’—〉5’方向移動。因而推測Rep蛋白和特定DNA解鏈酶是分別在DNA的兩條母鏈上協同作用以解開雙鏈DNA。
(3)DNA解鏈過程
DNA在複製前不僅為雙螺旋而且處於超螺旋狀態,而超螺旋狀態的存在為解鏈前的必須結構狀態,參與解鏈的除解鏈酶外有一些特定蛋白質,比如大腸桿菌中的Dna蛋白等。一旦DNA局部雙鏈被解開,就必須有ssbDNA蛋白以穩定解開單鏈,保證此局部不會恢復為雙鏈。兩條單鏈DNA複製的引發過程是有所差異,可是不論是前導鏈還是后隨鏈,都需要一段RNA引物用於開始子鏈DNA合成。因此前導鏈和后隨鏈的差別在於前者從複製起始點開始按5’—3’持續的合成下去、不形成岡崎片段、後者則隨著複製叉的出現、不斷合成長約2—3kb的岡崎片段。
岡崎片段與半不連續複製
因為DNA的兩條鏈是反向平行的,所以在複製叉附近解開的DNA鏈,一條為5’—〉3’方向,另一條為3’—〉5’方向,兩個模板極性是不同。所有已知DNA聚合酶合成方向均為5’—〉3’方向,不為3’—〉5’方向,所以無法解釋DNA的兩條鏈同時進行複製的問題。解釋DNA兩條鏈各自模板合成子鏈等速複製現象,日本的學者岡崎(Okazaki)等人提出了DNA的半不連續複製(semidiscontinuous replication)模型。在1968年,岡崎用3H脫氧胸苷短時間標記大腸桿菌,提取DNA,變性之後用超離心方法得到了許多3H標記的,被後人稱作為岡崎片段的DNA。延長標記時間之後,岡崎片段可轉變為成熟的DNA鏈,所以這些片段必然是複製過程中的中間產物。另一個實驗也證明DNA複製過程里首先合成較小的片段,即用DNA連接酶溫度敏感突變株進行的試驗,在連接酶不起作用的溫度中,便產生大量小DNA片段積累,表明DNA複製過程里至少有一條鏈首先合成較短的片段,之後再由連接酶鏈成大分子DNA。一般說,原核生物的岡崎片段比真核生物長。深入研究還可證明,前導鏈的連續複製與滯后鏈的不連續複製在生物界具有普遍性,故稱為DNA雙螺旋的半不連續複製。
在1941年,美籍印度人麥克林托克(Mc Clintock)就提出端粒(telomere)的假說,指出染色體末端必然存在一種特殊結構——端粒。已知染色體端粒的作用至少有2:a.保護染色體末端免受損傷,使染色體保持穩定;b. 與核纖層相連,使染色體得以定位。
弄清楚DNA複製過程之後,在20世紀70年代,科學家對DNA複製時新鏈5’端的RNA引物被切除之後,空缺為如何被填補的提出了質疑。如果不填補豈不是DNA每複製一次就短一點。后隨鏈複製為例,RNA引物被切除后,岡崎片段之間是由DNA聚合酶I催化合成的DNA填補之,然後再由DNA連接酶將它們連接成了一條完整的鏈。可是DNA聚合酶I催化合成DNA時,需要自由3’—OH作為其引物,最後餘下子鏈的5’則無法填補,於是染色體就短一點。
在正常體細胞里普遍存在著染色體酶複製一次端粒就短一次的現象。推測,可能一旦端粒縮短至某一閾限長度一下時,就會發出一個警報,指令細胞進入到衰老;或許為當細胞判斷出它們的染色體已變得太短了,所以是分裂也就停止了,造成了正常體細胞壽命有一定界限。可是在癌細胞中染色體端粒卻一直維持在一定長度上,這是為什麼,這是因為DNA複製之後,將染色體末端短缺部分補上需要端粒酶,是一種含有RNA的酶,其既解決了模板,又解決引物的問題。在生殖細胞與85%癌細胞中都測出了端粒酶具有活性,可是在正常體細胞中卻無活性,20世紀90年代中期Blackburn首次在原生動物中克隆出端粒酶基因。
端粒酶在癌細胞里具有活性,不僅使癌細胞可以不斷分裂增生,且為癌變前的細胞或已經是癌性的細胞提供了時間,積累附加的突變,即等於增加了它們複製,侵入與最終轉移的能力。同時人們也由此萌生開發以端粒為靶的藥物,即通過抑制癌細胞里端粒酶活性而達到治療癌症的目的。
至於真核細胞DNA末端結構特點,早就在1978年,Blackburn就以原生動物四膜出(一種纖毛蟲)為例說明之:a.迥紋形式的髮夾環;b.僅由C,A組成的簡單序列大量重複(C4A2)20~70;c.鏈上有許多缺口(nicks)
| 酶和蛋白質 | 作用 |
| 拓撲異構酶 | 幫助解開複製叉前後的超螺旋結構 |
| DNA解旋酶 | 解開螺旋 |
| Rep蛋白 | 幫助解開雙螺旋結構 |
| 引物合成酶 | 催化RNA引物合成並與DNA鏈互補的反應 |
| 單鏈結合蛋白 | 穩定單連區 |
| DNA聚合酶Ⅰ | 消除引物,填滿裂縫 |
| DNA聚合酶Ⅲ | 合成DNA |
| DNA連接酶 | 連接DNA末端 |
| RNA聚合酶 | 沿DNA模板轉錄一短的RNA分子 |
DNA新生鏈的合成由DNA聚合酶Ⅲ所催化,然而,DNA必須由螺旋酶在複製叉處邊移動邊解開雙鏈。這樣就產生了一種拓撲學上的問題:由於DNA的解鏈,在DNA雙鏈區勢必產生正超螺旋,在環狀DNA中更為明顯,當達到一定程度后就會造成複製叉難再繼續前進,從而終
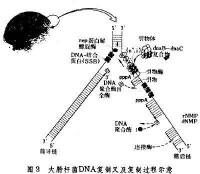
DNA複製
在DNA複製叉處要能由兩套DNA聚合酶Ⅲ在同一時間分別進行複製DNA前導鏈和滯后鏈。如果滯后鏈模板環繞DNA聚合酶Ⅲ全酶,並通過DNA聚合酶Ⅲ,然後再折向與未解鏈的雙鏈DNA在同一方向上,則滯后鏈的合成可以和前導鏈的合成在同一方向上進行。
這樣,當DNA聚合酶Ⅲ沿著滯后鏈模板移動時,由特異的引物酶催化合成的RNA引物即可以由DNA聚合酶Ⅲ所延伸。當合成的DNA鏈到達前一次合成的岡崎片段的位置時,滯后鏈模板及剛合成的岡崎片斷便從DNA聚合酶Ⅲ上釋放出來。這時,由於複製叉繼續向前運動,便產生了又一段單鏈的滯后鏈模板,它重新環繞DNA聚合酶Ⅲ全酶,並通過DNA聚合酶Ⅲ開始合成新的滯后鏈岡崎片段。通過這樣的機制,前導鏈的合成不會超過滯后鏈太多(最後只有一個岡崎片段的長度)。而且,這樣引發體在DNA鏈上和DNA聚合酶Ⅲ以同一速度移動。
多種DNA聚合酶在DNA複製過程中扮演不同的角色。在大腸桿菌中,DNA Pol III是主要負責DNA複製的聚合酶。它在複製分支上組裝成複製複合體,具有極高的持續性,在整個複製周期中保持完整。相反,DNA Pol I是負責用DNA替換RNA引物的酶。 DNA Pol I除了具有聚合酶活性外,還具有5'至3'外切核酸酶活性,並利用其外切核酸酶活性降解RNA引物。 Pol I在DNA複製中的主要功能是創建許多短DNA片段,而不是產生非常長的片段。在真核生物中,Pol α有助於啟動複製,因為它與引物酶形成複合物。Pol ε和Pol δ負責前導鏈的合成。Pol δ還負責引物的去除,而Pol ε也參與複製期間DNA的修復
按上述DNA複製的機制,在複製叉附近,形成了以兩套DNA聚合酶Ⅲ全酶分子、引發體和螺旋構成的類似核糖體大小的複合體,稱為DNA複製體(replisome)。複製體在DNA前導鏈模板和滯后鏈模板上移動時便合成了連續的DNA前導鏈和由許多岡崎片段組成的滯后鏈。在DNA合成延伸過程中主要是DNA聚合酶Ⅲ的作用。當岡崎片段形成后,DNA聚合酶Ⅰ通過其5'→3'外切酶活性切除岡崎片段上的RNA引物,同時,利用后一個岡崎片段作為引物由5'→3'合成DNA。最後兩個岡崎片段由DNA連接酶將其接起來,形成完整的DNA滯后鏈。
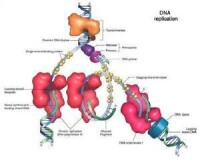
DNA複製
才終止其DNA複製。但最近的實驗表明,在DNA上也存在著複製終止位點,DNA複製將在複製終止位點處終止,並不一定等全部DNA合成完畢。但對複製終止位點的結構和功能了解甚少。在DNA複製終止階段令人困惑的一個問題是,線性DNA分子兩端是如何完成其複製的?已知DNA複製都要有RNA引物參與。當RNA引物被切除后,中間所遺留的間隙由DNA聚合Ⅰ所填充。但是,在線性分子的兩端以5'→3'為模板的滯后鏈的合成,其末端的RNA引物被切除后是無法被DNA聚合酶所填充的。
在研究T7DNA複製時,這個問題部分地得到了解決。T7DNA兩端的DNA序列區有160bp長的序列完全相同。而且,在T7DNA複製時,產生的子代DNA分子不是一個單位T7DNA長度,而是許多單位長度的T7DNA首尾連接在一起。T7DNA兩個子代DNA分子都會有一個3'端單鏈尾巴,兩個子代DNA的3'端尾巴以互補結合形成兩個單位T7DNA的線性連接。然後由DNA聚合酶Ⅰ填充和DNA連接酶連接后,繼續複製便形成四個單位長度的T7DNA分子。這樣複製下去,便可形成多個單位長度的T7DNA分子。這樣的T7DNA分子可以被特異的內切酶切開,用DNA聚合酶填充與親代DNA完全一樣的雙鏈T7DNA分子。
DNA複製
真核生物在染色體的多個點開始DNA複製,因此複製叉在染色體的許多點處相遇並終止。由於真核生物具有線性染色體,DNA複製無法到達染色體的最末端。由於這個問題,在染色體末端的DNA在每個複製周期中都會丟失。端粒是接近末端的重複DNA區域,有助於防止基因丟失。端粒縮短是體細胞中的正常過程,它縮短了子DNA染色體的端粒。因此,在DNA丟失阻止進一步分裂之前,細胞只能分裂一定次數。在生殖細胞中,端粒酶延伸端粒區域的重複序列以防止降解。
DNA複製的終止發生在特定的基因位點,即複製終止位點。該位點的終止位點序列被與該序列結合的阻止DNA複製的蛋白質識別並結合,阻止了複製叉前進,複製終止。細菌物的DNA複製末端位點結合蛋白又稱Ter蛋白。
因為細菌具有環狀染色體,所以當兩個複製叉在親本染色體的另一端彼此相遇時複製終止發生。大腸桿菌通過使用終止序列來調節該過程,當該序列被Tus蛋白結合時,終止序列僅允許複製叉一個方向的通行。結果,複製叉總是在染色體的終止區域內相遇,導致複製終止
在環狀DNA的複製的末端終止階段則不存在上述問題。環狀DNA複製到最後,由DNA拓撲異構酶Ⅱ切開雙鏈DNA,將兩個DNA分子分開成為兩個完整的與親代DNA分子一樣的子代DNA。
DNA複製的特點
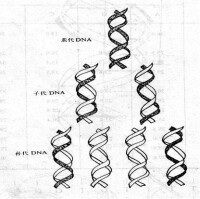
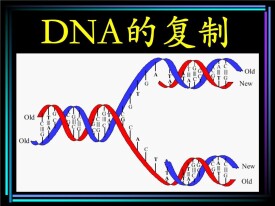
1.半保留複製:DNA在複製時,以親代DNA的每一股作模板,合成完全相同的兩個雙鏈子代DNA,每個子代DNA中都含有一股親代DNA鏈,這種現象稱為DNA的半保留複製。DNA以半保留方式進行複製,是在1958年由M. Meselson 和 F. Stahl 所完成的實驗所證明。
2.有一定的複製起始點:DNA在複製時,需在特定的位點起始,這是一些具有特定核苷酸排列順序的片段,即複製起始點(複製子)。在原核生物中,複製起始點通常為一個,而在真核生物中則為多個。
3.需要引物(primer):DNA聚合酶必須以一段具有3'端自由羥基(3'-OH)的RNA作為引物,才能開始聚合子代DNA鏈。RNA引物的大小,在原核生物中通常為50~100個核苷酸,而在真核生物中約為10個核苷酸。
4.雙向複製:DNA複製時,以複製起始點為中心,向兩個方向進行複製。但在低等生物中,也可進行單向複製。
5.半不連續複製:由於DNA聚合酶只能以5'→3'方向聚合子代DNA鏈,因此兩條親代DNA鏈作為模板聚合子代DNA鏈時的方式是不同的。以3'→5'方向的親代DNA鏈作模板的子代鏈在聚合時基本上是連續進行的,這一條鏈被稱為領頭鏈(leading strand)。而以5'→3'方向的親代DNA鏈為模板的子代鏈在聚合時則是不連續的,這條鏈被稱為隨從鏈(lagging strand)。DNA在複製時,由隨從鏈所形成的一些子代DNA短鏈稱為岡崎片段(Okazaki fragment)。岡崎片段的大小,在原核生物中約為1000~2000個核苷酸,而在真核生物中約為100個核苷酸。
DNA的複製是一個邊解旋邊複製的過程。複製開始時,DNA分子首先利用細胞提供的能量,在解旋酶的作用下,把兩條螺旋的雙鏈解開,這個過程叫解旋。然後,以解開的每一段母鏈為模板,以周圍環境中的四種脫氧核苷酸為原料,按照鹼基配對互補配對原則,在DNA聚合酶的作用下,各自合成與母鏈互補的一段子鏈。隨著解旋過程的進行,新合成的子鏈也不斷地延伸,同時,每條子鏈與其母鏈盤繞成雙螺旋結構,從而各形成一個新的DNA分子。這樣,複製結束后,一個DNA分子,通過細胞分裂分配到兩個子細胞中去!
註:複製時遵循鹼基互補配對原則,複製發生在細胞分裂的間期。
DNA是遺傳信息的載體,故親代DNA必須以自身分子為模板準確的複製成兩個拷貝,並分配到兩個子細胞中去,完成其遺傳信息載體的使命。而DNA的雙鏈結構對於維持這類遺傳物質的穩定性和複製的準確性都是極為重要的。
(一)DNA的半保留複製
Waston和Click在提出DNA雙螺旋結構模型時曾就DNA複製過程進行過研究,他們推測,DNA在複製過程中鹼基間的氫鍵首先斷裂,雙螺旋解旋分開,每條鏈分別作模板合成新鏈,每個子代DNA的一條鏈來自親代,另一條則是新合成的,故稱之為半保留式複製(semiconservative replication)。
1958年Meselson和Stahl進行了如圖8-3-5的實驗證明了DNA分子是以半保留方式進行自我複製的。圖8-3-5 Meselson和Stahl證明DNA半保留複製的實驗
(二)DNA複製的起始,方向和速度
DNA在複製時,雙鏈DNA解旋成兩股分別進行。其複製過程的複製起點呈現叉子的形式,故稱複製叉。以複製叉向前移動的方向為標準,一條模板鏈為3’—〉5’走向,在其上DNA能以5’—〉3’方向連續合成,稱為前導鏈(leading strand);另一條模板鏈為5’—〉3’走向,在其上DNA也是5’—〉3’方向合成,但與複製叉移動的方向正好相反,故隨著複製叉的移動形成許多不連續的岡崎片段,最後在連成一條完整的DNA鏈,該鏈稱為後隨鏈(lagging strand)。實驗證明DNA的複製是由一個固定的起始點開始的。一般把生物體的單個複製單位稱為複製子。一個複製子只含一個複製起點。一般說,細菌,病毒即線粒體DNA分子均作為單個複製子完成其複製,真核生物基因組可以同時在多個複製起點上進行雙向複製,即它們的基因組包括多個複製子。多方面的實驗結果表明,大多數生物內DNA的複製都是從固定的起始點以雙向等速方式進行的。複製叉以DNA分子上某一特定順序為起始點,向兩個方向等速生長前進。圖8-3-6 DNA複製過程
DNA複製過程
(三)DNA複製過程
以原核生物DNA複製過程予以簡要說明
1.DNA雙螺旋的解旋
DNA在複製時,其雙鏈首先解開,形成複製叉,而複製叉的形成則是由多種蛋白質及酶參與的較複雜的複製過程
ssbDNA蛋白是較牢固的結合在單鏈DNA上的蛋白質。原核生物ssbDNA蛋白與DNA結合時表現出協同效應:若第1個ssbDNA蛋白結合到DNA上去能力為1,第2個的結合能力可高達103;真核生物細胞中的ssbDNA蛋白與單鏈DNA結合時則不表現上述效應。ssbDNA蛋白的作用是保證解旋酶解開的單鏈在複製完成前能保持單鏈結構,它以四聚體的形式存在於複製叉處,待單鏈複製后才脫下來,重新循環。所以,ssbDNA蛋白只保持單鏈的存在,不起解旋作用。(2)DNA解鏈酶(DNA helicase) DNA解鏈酶能通過水解ATP獲得能量以解開雙鏈DNA。這種解鏈酶分解ATP的活性依賴於單鏈DNA的存在。如果雙鏈DNA中有單鏈末端或切口,則DNA解鏈酶可以首先結合在這一部分,然後逐步向雙鏈方向移動。複製時,大部分DNA解旋酶可沿滯后模板的5’—〉3’方向並隨著複製叉的前進而移動,只有個別解旋酶(Rep蛋白)是沿著3’—〉5’方向移動的。故推測Rep蛋白和特定DNA解鏈酶是分別在DNA的兩條母鏈上協同作用以解開雙鏈DNA。(3)DNA解鏈過程 DNA在複製前不僅是雙螺旋而且處於超螺旋狀態,而超螺旋狀態的存在是解鏈前的必須結構狀態,參與解鏈的除解鏈酶外還有一些特定蛋白質,如大腸桿菌中的Dna蛋白等。一旦DNA局部雙鏈解開,就必須有ssbDNA蛋白以穩定解開的單鏈,保證此局部不會恢復成雙鏈。兩條單鏈DNA複製的引發過程有所差異,但是不論是前導鏈還是后隨鏈,都需要一段RNA引物用於開始子鏈DNA的合成。因此前導鏈與后隨鏈的差別在於前者從複製起始點開始按5’—3’持續的合成下去,不形成岡崎片段,後者則隨著複製叉的出現,不斷合成長約2—3kb的岡崎片段。
2.岡崎片段與半不連續複製
因DNA的兩條鏈是反向平行的,故在複製叉附近解開的DNA鏈,一條是5’—〉3’方向,另一條是3’—〉5’方向,兩個模板極性不同。所有已知DNA聚合酶合成方向均是5’—〉3’方向,不是3’—〉5’方向,因而無法解釋DNA的兩條鏈同時進行複製的問題。為解釋DNA兩條鏈各自模板合成子鏈等速複製現象,日本學者岡崎(Okazaki)等人提出了DNA的半連續複製(semidiscontinuous replication)模型。1968年岡崎用3H脫氧胸苷短時間標記大腸桿菌,提取DNA,變性後用超離心方法得到了許多3H標記的,被後人稱作岡崎片段的DNA。延長標記時間后,岡崎片段可轉變為成熟DNA鏈,因此這些片段必然是複製過程中的中間產物。另一個實驗也證明DNA複製過程中首先合成較小的片段,即用DNA連接酶溫度敏感突變株進行試驗,在連接酶不起作用的溫度下,便有大量小DNA片段積累,表明DNA複製過程中至少有一條鏈首先合成較短的片段,然後再由連接酶鏈成大分子DNA。一般說,原核生物的岡崎片段比真核生物的長。深入研究還證明,前導鏈的連續複製和滯后鏈的不連續複製在生物界具有普遍性,故稱為DNA雙螺旋的半不連續複製。
3.複製的引發和終止
所有的DNA的複製都是從一個固定的起始點開始的,而DNA聚合酶只能延長已存在的DNA鏈,不能從頭合成DNA鏈,新DNA的複製是如何形成的?經大量實驗研究證明,DNA複製時,往往先由RNA聚合酶在DNA模板上合成一段RNA引物,再由聚合酶從RNA引物3’端開始合成新的DNA鏈。對於前導鏈來說,這一引發過程比較簡單,只要有一段RNA引物,DNA聚合酶就能以此為起點,一直合成下去。對於后隨鏈,引發過程較為複雜,需要多種蛋白質和酶參與。后隨鏈的引發過程由引發體來完成。引發體由6種蛋白質構成,預引體或引體前體把這6種蛋白質結合在一起並和引發酶或引物過程酶進一步組裝形成引發體。引發體似火車頭一樣在後隨鏈分叉的方向前進,並在模板上斷斷續續的引發生成滯后鏈的引物RNA短鏈,再由DNA聚合酶 III 作用合成DNA,直至遇到下一個引物或岡崎片段為止。由RNA酶H降解RNA引物並由DNA聚合酶 I 將缺口補齊,再由DNA連接酶將每兩個岡崎片段連在一起形成大分子DNA.。
(四)端粒和端粒酶
1941年美籍印度人麥克林托克(Mc Clintock)就提出了端粒(telomere)的假說,認為染色體末端必然存在一種特殊結構——端粒。已知染色體端粒的作用至少有二:① 保護染色體末端免受損傷,使染色體保持穩定;② 與核纖層相連,使染色體得以定位。
在弄清楚DNA複製過程之後,20世紀70年代科學家對DNA複製時新鏈5’端的RNA引物被切除后,空缺是如何被填補的提出了質疑。如不填補豈不是DNA每複製一次就短一點。以後隨鏈複製為例,當RNA引物被切除后,岡崎片段之間是由DNA聚合酶 I催化合成的DNA填補之,然後再由DNA連接酶將它們連接成一條完整的鏈。但是DNA聚合酶 I 催化合成DNA時,需要自由3’—OH作為引物,最後餘下子鏈的5’無法填補,於是染色體就短了一點。
在正常體細胞中普遍存在著染色體酶複製一次端粒就短一次的現象。人們推測,可能一旦端粒縮短到某一閾限長度一下時,他們就會發出一個警報,指令細胞進入衰老;或許是當細胞判斷出它們的染色體已變得太短了,於是分裂也就停止了,造成正常體細胞壽命有一定界限。但是在癌細胞中染色體端粒卻一直維持在一定長度上,這是為什麼?這是因為DNA複製后,把染色體末端短缺部分補上需要端粒酶,這是一種含有RNA的酶,它既解決了模板,又解決了引物的問題。在生殖細胞和85%癌細胞中都測出了端粒酶具有活性,但是在正常體細胞中卻無活性,20世紀90年代中期,Blackburn首次在原生動物中克隆出端粒酶基因。
端粒酶在癌細胞中具有活性,它不僅使癌細胞可以不斷分裂增生,而且它為癌變前的細胞或已經是癌性的細胞提供了時間,以積累附加的突變,即等於增加它們複製,侵入和最終轉移的能力。同時人們也由此萌生了開發以端粒為靶的藥物,即通過抑制癌細胞中端粒酶活性而達到治療癌症的目的。
至於真核細胞DNA末端的結構特點,早就在1978年Blackburn就以原生動物四膜蟲(一種纖毛蟲)為例說明之:① 迥紋形式的髮夾環;② 僅由C,A組成的簡單序列大量重複(C4A2)20~70;③ 鏈上有許多缺口(nicks)。
基本信息
- 中文名
- DNA複製
- 外文名
- DNA replication
- 拉丁學名
- replication
- 途徑
- 半保留複製
- 簡寫
- DNA
- 作用
- 生物遺傳的基礎
- 載體
- 所有DNA為遺傳物質的生物體
- 過程
- 引發、延伸、終止