共找到2條詞條名為維管植物的結果 展開
- 維管植物
- 導管植物
維管植物
維管植物
維管植物(vascular plant)是具有維管組織的植物。現存的維管植物大約有25~30萬種,包括極少部分苔蘚植物、蕨類植物(松葉蘭類、石松類、木賊類、真蕨類)、裸子植物和被子植物。維管系統(木質部和韌皮部)的發生是植物從水生到陸生長期適應環境的結果。維管系統的有效輸導,使維管植物成為最繁茂的陸生植物。比較原始的維管植物的木質部中多隻具管胞,故也可稱這些植物為管胞植物。
凡是有維管系統(vascularsystem)的植物都稱維管植物,包括蕨類和種子植物;它們與藻類、菌類、地衣、苔蘚植物不同之處在於具有發達的維管系統;維管系統主要由木質部和韌皮部組成,木質部中含有運輸水分和無機鹽的管胞或導管分子,韌皮部中含有運輸糖類等的篩胞或篩管。它們大多為陸生植物,只有少數在受精過程需要在水中進行。它們的孢子體在生活史中佔優勢,只有在幼小的時候(即在胚胎階段)才依賴配子體而生存。它們的配子體一般較小,在松柏類和有花植物中其配子體完全寄生在孢子體中,孢子體的形態結構有的較簡單,僅為軸形,有的則分化成複雜的根、莖、葉系統。

維管植物
木質部由四種細胞組成:管胞、導管節、薄壁組織及纖維。韌皮部則由五種細胞組成:篩管節、伴細胞、薄壁組織、纖維及厚壁細胞。
水份在木質部由根部向上到植物的莖部及葉片,這逆地心吸力的過程主要由蒸騰作用維持。
由初生木質部和初生韌皮部所組成的維管組織是一種初生結構。它們聚集而成中柱(stele)。按照維管組織排列方式的不同而形成多種類型的中柱。根據中柱類型可以判斷植物類群之間的親緣關係。
中柱可以分為下列5種類型:(1)原生中柱(protostele),包括單中柱(haplostele)、星狀中柱(actinostele)和編織中柱(plectostele);(2)管狀中柱(siphonostele)包括雙韌管狀中柱(amphiphloicsiphonostele)和外韌管狀中柱(ectophloicsiphonostele);(3)網狀中柱(dictyostele);(4)真中柱(eustele);(5)散生中柱(atactostele)。茲將各種類型中柱的特點分敘如下。
(一)原生中柱
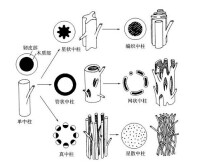
各種圍觀植物的中柱
(二)管狀中柱
管狀中柱的特點是木質部圍繞中央髓形成圓筒狀。若韌皮部在木質部的內外兩邊都出現則稱為雙韌管狀中柱有許多蕨類植物具有此種中柱。雙韌管狀中柱又稱疏隙中柱(solenostele)。若韌皮部位於木質部的外部表面則為外韌管狀中柱。管狀中柱在蕨類植物中普遍存在。
(三)網狀中柱
網狀中柱由管狀中柱演變而來。由於莖的節間甚短,節部位葉隙密集,從而使中柱產生許多裂隙,從橫剖面上看中柱被割成一束束。每一束中央為木質部,木質部外面圍著韌皮部,而韌皮部外再圍著內皮層。有不少蕨類植物具有此種類型的中柱。
(四)真中柱
真中柱。的木質部與韌皮部並列成束或索狀。蕨類中的木賊屬(Happochaete)及多數裸子植物,被子植物具此種類型的中柱。
(五)散生中柱
在單子葉植物中,維管組織分散於莖內。此種中柱稱散生中柱。
1963年,韋特蒙(Wetmon)和里爾(Rier)曾用實驗方法研究維管組織分化。將未分化的薄壁愈傷組織進行組織培養,發現若將與愈傷組織相同種的芽嫁接上去則導致在愈傷組織中產生維管組織的小結,但此種維管組織並不繼續發展。這就清楚地說明維管組織的分化是受化學物質所控制。他們的進一步研究表明生長激素和糖能局部地導致維管組織的出現。他們對糖(葡萄糖和蔗糖)的濃度也進行了測定,發現1.5—2.0%的蔗糖只導致產生木質部;若濃度提高到3—3.5%,則在愈傷組織中產生出中央為木質部,周圍為韌皮部的維管組織,在木質部與韌皮部之間並能產生形成層;濃度若再提高到4—4.5%則刺激產生韌皮部,使韌皮部逐漸佔優勢。若將生長激素與糖的混合物用毛細管注入愈傷組織,則導致形成筒狀木質部和韌皮部,兩者之間有形成層。此項研究充分說明了生長激素與糖在不同濃度情況下對愈傷組織分化所起的作用。
各種不同類型的中柱與植物的進化有何關係?對此有一種假說:若原生中柱中央的木質部被薄壁組織所取代則發展成筒狀中柱。此種假說的佐證是有時在髓中會出現木質部的成分(管胞),此一過程被稱為髓形成作用(medullation)。由於葉隙的大量出現,節間的縮短,便使管狀中柱演化成真中柱和散生中柱,亦即在種子植物中所見到的中柱的最高級的形式。
對維管植物的分類尚存在分歧。一種觀點認為維管植物有其共同祖先,它們是單元起源的(monophyletic),也就是說它只有一次起源,所有的維管植物都是由最初形成的原始祖先分化發展而來。因此,所有維管植物在分類系統中應歸成一門,即維管植物門(Tracheo-phyta)。但是具有維管組織的植物千差萬別,甚至在海藻中也發現了維管組織的分子,它與其他維管植物在系統演化上相差甚遠,無法歸成一類。

維管植物
維管植物具有孢子體世代和配子體世代。孢子體非常發達,具有根、莖、葉等營養器官,並能產生具孢子的孢子囊。低等維管植物的配子體可以獨立生活,但是形狀細小,結構簡單。到了種子植物,配子體更為退化,只有幾個細胞在孢子內發育。維管植物的孢子體可以由有性生殖的合子或無性生殖的植物體的一部分或單個細胞形成。合子經過不斷的細胞分裂,形成胚,從中分化出根端分生組織與莖端分生組織(見頂端分生組織)並建立起體軸系統。由胚發育成幼苗,並分化出各種組織系統。孢子體世代產生或多或少的孢子囊,每個孢子囊中可產生幾個或多個孢子。產生孢子時,都經過細胞的減數分裂,染色體減半。由孢子中發育出配子體,從中形成頸卵器與精子器,或者形成更簡化的樣式。配子體中產生卵(雌配子)與精子(雄配子)。卵與精子結合成為合子,從而又開始新的孢子體世代。
維管植物主要的辨別方法:維管植物具有維管組織,可以讓植物生長到一個較大的體積。非維管植物則一直保持著較小的體積。維管植物主要生成階段是孢子體。在木質部和韌皮部,水分皆會被不停運送:木質部將水和無機溶質從根部運送往葉片,而韌皮部則會把植物中的有機溶質送往全株植物。維管植物都含有木質化的組織(即木質導管或管胞) 。
維管植物光合作用的產物是甲烷產生的主要碳源,植物根系的周轉和碳物質的分泌為產甲烷細菌提供底物;維管植物根際氧化是甲烷氧化最主要的途徑,在植物的生長期佔到總氧化量的80%左右。植物傳輸O2的能力和根際O2的需求是影響根際氧化的主要因素;維管植物通氣組織的傳輸促進了甲烷從土壤向大氣的輸送,但所採用的傳輸機制影響著氣體的輸送效率。此外,自然濕地甲烷排放的各個過程均受到維管植物形態和植被類型的影響。維管植物在甲烷排放中的作用,可以部分解釋自然濕地甲烷在排放的時間(季節性變化、日變化)和空間尺度上的差異。維管植物對於自然濕地甲烷排放具有指示意義,可以用於大尺度自然濕地甲烷排放量的估算
濕地甲烷排放過程受到許多物理, 化學, 生物因素的共同影響, 單因子關係不足以解釋甲烷排放的差異。通過分析, 在濕地分佈廣泛的維管植物對自然濕地甲烷排放的產生、氧化、傳輸過程都有重要的影響。由於植物對底質的生物地化循環既有直接影響, 還有間接影響, 並且這些過程緊密相連。加之研究方法不足, 區分和定量這些過程依然是機制研究的難點。特別是控制甲烷排放各個過程的微生物群落和植物之間的相互作用, 有待於進一步的研究。在區域或全球尺度自然濕地甲烷排放量估算方面, 需要進行更多尤其是不同植物類型的通量觀測, 需要對遙感數據中植被覆蓋信息進行更精確的提取, 需要發展更完善的綜合氣候、環境和植被因素的生物過程模型。

基本信息
- 中文名
- 維管植物
- 外文名
- vascular plant
- 別名
- 管胞植物
- 界
- 體征
- 由初生木質部和初生韌皮部所組成的維管組織是一種初生結構
- 電子產品類別
- 25~30萬種
- 包含種類
- 蕨類植物和種子植物
- 繁殖方式
- 孢子體和配子體世代繁殖