秀麗隱桿線蟲
秀麗隱桿線蟲
秀麗隱桿線蟲(學名:Caenorhabditis elegans),是一種可以獨立生存的線蟲,長度約1mm,生活在溫度恆定的環境。秀麗隱桿線蟲具有極高的科學研究價值,常被作為分子生物學和發育生物學研究領域的模式生物。
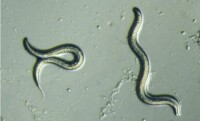
秀麗隠桿線蟲
秀麗隠桿線蟲有雄性及雌雄同體兩種性別,其基本解剖構造包括一個口、咽、腸、性腺,及膠原蛋白角質層。雄性有個單葉的性腺、輸精管及一個特化為交配用的尾部。雌雄同體有兩個卵巢、輸卵管、藏精器,及單一的一個子宮。
從研究的角度來看,秀麗隠桿線蟲的優勢在於為一種多細胞真核生物,又夠簡單到可以被詳細研究。秀麗隠桿線蟲每一個體細胞(雌雄同體成蟲有959個;雄成蟲有1031個)的發展命運都已被確立。這個細胞世系的規律在各個個體之間是幾乎不變的。兩種性別的個體,都有許多多出的細胞(雌雄同體131個,大部分原本將成為神經元)會經由細胞凋亡的過程被除去。
野生型秀麗隠桿線蟲雌雄同體,每一細胞之細胞核均被染色顯示。除此之外,秀麗隠桿線蟲還是神經系統最簡單的生物之一。在雌雄同體中,總共有302個神經元,其連結形式也已完全被建立出來,且被證實為一小世界網路。更多研究探索了與秀麗隠桿線蟲的一些特殊行為有密切關連的神經機制,包括化學趨向性、趨溫性,以及雄性交配行為。
2002年,諾貝爾生理醫學獎頒發給了Sydney Brenner與H. Robert Horvitz和John Sulston,以表揚他們對了解秀麗隠桿線蟲細胞死亡程序及器官發育遺傳學的貢獻。

好
秀麗隠桿線蟲在實驗室中20°C的情況下,平均壽命約為二、三周,而發育時間只須幾天。
胚胎髮生可以大致分成兩個時期:增殖期和器官與型態形成期。在增值期受精卵會從一個細胞逐漸增殖成大約550個必要的未分化細胞,而增殖期又可以分為兩個階段,其中一個階段是在母體內進行(在22°C的生長環境下約為受精后0-150分鐘),這個階段分裂出較少的創始者細胞,在增殖期結束時,胚胎形成一個含有三胚層的球型構造,這三個胚層分別是外胚層(之後分化生成皮下組織和神經系統)、中胚層(未來產生咽部和肌肉系統)和內胚層(以後生成生殖腺和腸道)。而另一個階段則進行大量的細胞分裂和原腸形成(在22°C的生長環境下約為受精后150-350分鐘),這個階段持續到胚胎進入器官與型態形成期。
在22°C的生長環境下約為受精后的5.5-5.6小時到12-14小時,胚胎會增長約三倍並形成完全分化的組織和器官,根據胚胎內觀察到的蟲體摺疊數可以分為comma stage、1.5摺疊期、2摺疊期、3摺疊期以及4摺疊期。第一次的肌肉抽動在受精后430分鐘可以被觀察到(約為1.5摺疊或2摺疊期);而在受精后約510分鐘不同性別可以觀察到發育差異(雌雄間性胚胎的頭部伴護神經(cephalic companion neurons)死亡,雄性胚胎的雌雄間性特有神經(hermaphrodite-specific neurons)死亡),而在3摺疊期的晚期;蟲體運動神經系統已發育且可以在蛋裡頭順著其長軸進行移動;而在4摺疊期時(第一次細胞分裂后760分鐘)胚胎的咽部開始進行收縮抽動;而在第一次細胞分裂后800分鐘由蛋中孵化。
后胚胎髮育期由孵化後有食物提供刺激下啟動,在有食物的情況下,細胞分裂持續且后胚胎髮育開始於孵化后的三個小時,一般而言秀麗隱桿線蟲經歷四個階段的幼蟲期(L1、L2、L3、L4)后變成成體,許多在胚胎期即設置好的胚細胞於這四個階段的幼蟲期以時間及空間規劃幾乎不變的模式進行分裂,而這也給予了秀麗隱桿線蟲固定數量細胞以及命定的細胞命運。而在胚胎期所產生的671個細胞核中,其中有113個細胞會在後胚胎髮育的過程會進行計劃性細胞凋亡,而剩下的558個細胞中只有百分之十的細胞(雌雄間性有51個細胞,雄性有55個細胞)是胚細胞且可以再進一步進行分裂。
秀麗隠桿線蟲
在L2幼蟲期的末期,如果環境狀況不適合繼續生長的話,秀麗隱桿線蟲的幼蟲可能會進入dauer幼蟲階段,這些不適合生長的環境狀況包括受到環境費洛蒙影響、食物匱乏、高溫等,會促使幼蟲進入L2d的幼蟲階段,這個階段幼蟲同時具有可以繼續進入L3幼蟲期或dauer幼蟲期的潛力,若環境逆境太強則進入dauer,若環境轉好則進入L3幼蟲期,而dauer期為一個不會衰老的狀態,因為dauer期的長短並不會影響dauer期后的蟲體壽命。在取得食物供應后的一個小時內,dauer幼蟲脫離dauer,二到三個小時之後開始進食,最後在十個小時後會蛻皮進入L4幼蟲期。
在22°C到25°C的環境下,大概在孵化后的45-50小時就會變成成熟的雌雄間性併產下他的第一個卵,成體雌雄間性大約可以產生四天的卵細胞,在這段具有生殖力期間后的三到四天,成體會另外再存活十到十五天,自體受精的雌雄間性會產生大約300個後代(因為精子的數目是有限的),但是如果和雄性交配的話,雌雄間性會優先採用雄性的精子,此時後代數目可以增加到1200-1400個。雄性則在最後一次幼蟲蛻皮后的六天具有和雌雄間性交配的能力,且大概可以產生3000個後代。
1、秀麗隠桿線蟲以微生物為食,如大腸桿菌(Escherichia coli),易於大量培養,每隻成蟲在生命周期里可產生約300隻後代,適合作遺傳學研究。
2、培養秀麗隱桿線蟲成本低,在實驗室中容易掌控,並且可以在不用時冷凍,解凍之後仍能存活,因此適合長時間儲存。
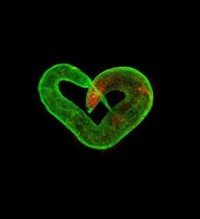
秀麗隠桿線蟲
4、秀麗隱桿線蟲神經系統相對簡單,於雌雄同體之中,有302個神經元,約7000個突觸。悉尼·布倫納等人將秀麗隱桿線蟲續列縱切,利用電子顯微鏡得到的影像,重建其神經系統,並在1986年發表雌雄同體各個神經元之間如何連結,成為世界上第一個,也是目前唯一的神經網路體(英語:connectome)。2012年雄性的身體後部神經系統也被重建,助於科學家了解交配行為所需的神經系統結構。許多神經生物學家在此之後,利用秀麗隱桿線蟲做為模式生物,來了解其如何產生趨化性,趨溫性,力傳導及交配行為。
自1965年起,科學家悉尼·布倫納利用線蟲作為分子生物學和發育生物學研究領域的模式生物。1974年悉尼·布倫納提出以秀麗隱桿線蟲做為模式生物,以研究發育生物學及神經科學的重要課題。

被帶去太空的線蟲
2009年11月17日,美國宇航局的“阿特蘭蒂斯號”太空梭將百萬秀麗隱桿線蟲送入了太空,旨在幫助諾丁漢大學的專家進一步了解什麼因素引起人體肌肉增強和萎縮。在空間站上,這些線蟲將經歷導致宇航員肌肉出現戲劇性萎縮的無重力狀態。
瑟奇亞克將為其中一些秀麗隱桿線蟲注入一種名為“RNAi”的物質,這種物質能夠幫助降低肌肉消耗,它們的目的地空間站距離地球大約200英里(約合322公里)。為了這次飛行,這些精挑細選的線蟲已進入一種睡眠狀態。它們被放入特製的細胞培養袋並進入太空,而後在食物的幫助下從睡眠中蘇醒過來。進入太空之後,它們將暴露在零重力環境下,時間為4天,而後接受冷凍,為重返地球做準備。回到諾丁漢大學的實驗室之後,科學家將對此次太空飛行對肌肉質量產生何種影響進行分析。

在顯微鏡下很容易看到一段藍色熒光穿過蠕蟲內臟時的死亡蔓延。這種熒光由細胞壞死造成,它在有機體內的傳播依賴於鈣傳遞信號。
在研究中,科學家抑制影響胰島素活性和營養信號通路——雷帕黴素標靶的關鍵分子。雷帕黴素標靶通路的單一基因突變使秀麗隱桿線蟲壽命延長30%,但胰島素信號傳遞突變使它們的壽命增加一倍。如果同時使用這兩個方面,它們的壽命可能延長130%,但它們結合起來的效果更大。
細胞凋亡現象及其機理,最早是在線蟲中被揭示的。凋亡(apoptosis)是一個希臘文來源的詞語,這個字眼表達的是花兒凋謝,樹葉飄零的景色。“梧桐一葉落而知天下秋”、“搦搦兮秋風,洞庭波兮木葉下”的意象恐怕正是說的這種意境:優雅,含蓄,還帶點淡淡的憂傷,更因為飄落時那種雖然有些無奈卻坦然以受之的美。由於線蟲研究開創了一個對今日生物醫學發展,具有舉足輕重的全新領域,同時也因為以線蟲為基礎的凋亡研究對基礎和應用生物學,產生的巨大推動作用,卡羅林斯卡醫學院的諾貝爾獎評選委員,會將年2002年生理和醫學獎授予了,線蟲生物學的開拓者:西德尼·布雷納(Sydney Brenner)、約翰·薩爾斯頓(John Sulston)和線蟲凋亡之父羅伯特·霍維茨(Robert Horvitz)。
今天,秀麗線蟲因其遺傳背景清楚、個體結構簡單、生活史短、基因組測序完成等,在遺傳與發育生物學、行為與神經生物學、衰老與壽命、人類遺傳性疾病、病原體與生物機體的相互作用、藥物篩選、動物的應急反應、環境生物學和信號傳導等領域得到廣泛應用。通過各國科學家的密切合作,線蟲研究的資源共享體系,為研究人員提供了極大的方便,如由NIH資助的線蟲種質中心(CGC)貯藏了大量的線蟲品系,免費分發給世界各地的研究者,而AceDB系統為研究者提供了大量的有關線蟲的信息,線蟲的DNA序列可以在NCBI或Wormbase網站上進行在線比對。
1900年,Maupas把這種棒狀的蠕蟲命名為Rhabditis elegans,因為群體繁殖為“r-選擇”的緣故,早在1948年Dougherty and Calhoun 便指出了它在遺傳學研究中的重要性。1952年,Osche把它置於Caenorhabditis亞屬,1955年Dougherty最終把它命名為Caenorhabditis elegans(其中Caeno意為 recent; rhabditis意為 rod; elegans意為nice)。廣泛使用N2 Bristol品系,由Staniland從英格蘭Bristol附近的蘑菇堆肥中分離,1965年被定為參考種N2。在此過程中,Dougherty建立了線蟲的瓊脂板接種,大腸桿菌的培養方法和無菌單培養方法。對秀麗線蟲的早期研究工作,主要集中在解剖、營養、生理和生殖等方面,直到1960s中期,隨著DNA雙螺旋結構的揭示和遺傳密碼的發現,使得當時的生物學家認為,“人腦是生命科學研究的最後堡壘。”
1963年,Sydney Brenner在寫給劍橋大學分子生物學實驗室主任Perutz的信中指出:“幾乎所有的經典的分子生物學問題已經解決或在今後幾十年內將得到解決,……今後分子生物學的任務,是深入的探討生命科學的其它領域,特別是發育和神經系統”。在此背景下,Brenner著手尋找一種具有生活周期短,個體小容易繁殖的後生動物,1963年10月,他從Dougherty實驗室獲得了線蟲Bristol品系,並對此開展不懈的研究。在早期的研究工作中,Brenner分離得到了一些營養突變體,他轉而使用EMS處理線蟲,首先獲得一種短小矮胖的個體,被命名為dpy-1。
到1974年Brenner的論文The Genetics of Caenorhabditis elegans發表時,通過EMS誘變一共獲得300多個線蟲突變體,其中多數為隱性突變。突變表型涉及行為、運動和形態結構等方面,這些突變材料使得100多個基因得以表徵,並被定位於6個連鎖群。秀麗線蟲作為模式生物的優越性也日漸顯示,如通體透明,體細胞數目恆定,特定細胞位置固定等,這使得它成為研究個體發生的良好材料。Sulston通過活體觀察線蟲的胚胎髮育和細胞遷移途徑,於1983年完成線蟲從受精卵到成體的細胞譜系,是發育生物學史上具有里程碑性的發現,隨後秀麗線蟲在胚胎髮育、性別決定、細胞凋亡、行為與神經生物學等方面研究中得到廣泛應用。其中Brenner,Sulston和Horvitz因在線蟲的遺傳與發育方面的成就,獲得2002年諾貝爾醫學或生理學獎。從1980年代中期開始的線蟲基因組測序工作,於1998年完成,同年Fire建立了線蟲RNA干擾技術,該技術可以方便的沉默特定的基因,即通過反向遺傳學(reverse genetics)研究特定基因的功能,在生命科學的許多領域得到應用,2006年Fire和Mello因此而獲得諾貝爾醫學或生理學獎。
基本信息
- 中文名
- 秀麗隠桿線蟲
- 別名
- 秀麗新小桿線蟲、秀麗線蟲
- 拉丁學名
- Caenorhabditis elegans
- 界
- 動物界
- 門
- 線蟲動物門
- 綱
- 線蟲綱
- 亞綱
- 小桿亞綱
- 目
- 小桿線蟲目
- 科
- 小桿科
- 屬
- 廣桿線蟲屬
- 亞屬
- Caenorhabditis
- 種
- 秀麗隱桿線蟲
- 分布區域
- 全球廣泛分佈
- 命名者及年代
- C. elegans
- 體征
- 有口有肛門