子囊菌亞門真菌
子囊菌亞門真菌
子囊菌亞門真菌一般稱作子囊菌,將近有15000種,它們的形態、生活史和生活習性的差別很大,共同的特徵是有性生殖形成子囊孢子。子囊菌大都是陸生的,有腐生和寄生的,不少是植物病原物。子囊菌的營養體是很發達的、有隔膜的菌絲體,少數(如酵母菌)是單細胞的。許多子囊菌的菌絲體可以形成菌組織,因而有子座和菌核等機構。無性繁殖產生分生孢子,有些子囊菌經常產生的是分生孢子(conidia)。由於分生孢子的形成在許多子囊菌的生活史中占很重要的位置,所以它的無性階段也稱作分生孢子階段。當然,子囊菌也有不產生分生孢子的。關於分生孢子梗和分生孢子的各種形狀以及它們的著生方式,將在半知菌亞門真菌中說明。
是從菌絲體上分別形成多核的、較小的雄器和較大的產囊體,產囊體上有授精絲。當雄器與授精絲接觸后,在接觸點形成一個孔口,雄器中的許多細胞核就通過授精絲進入產囊體與其中的許多細胞核交配形成成對的雙核。交配后,從產囊體上形成若干產囊絲,產囊絲還可以分枝。產囊絲和它的分枝頂端細胞有一對雙核,發育而形成子囊和子囊孢子。事實上,許多子囊菌的雄器和產囊體的分化程度並不這樣明顯,它們也可以都是單核的,也有不形成雄器而是以很小的性細胞與產囊體交配。
各種子囊菌交配(質配)的形式不同
但是從產囊絲形成子囊和子囊孢子的過程是大致相似的(圖2-24)。產囊絲和分枝的頂端細胞,先彎曲成鉤狀體。鉤狀體中的雙核並裂后形成兩個隔膜,分隔為3個細胞,頂端和基部細胞都是單核的,中間雙核的稱作子囊母細胞。子囊母細胞中的雙核進行核配成為一個二倍體的細胞核。子囊母細胞伸長,其中二倍體的細胞核進行減數分裂,又經過一次有絲分裂,最後形成8個單倍體的細胞核。這些細胞核和它們周圍的細胞質形成8個子囊孢子。在形成子囊孢子時,子囊中還有部分細胞質的殘餘物,其中含有肝糖。當子囊孢子成熟時,由於肝糖的水解而提高了子囊的滲透壓,當壓力超過子囊壁的彈性,子囊的頂部就開裂而將子囊孢子彈出。
一些較低等的子囊菌,有性生殖的方式則與此完全不同。它們並不形成特殊的配子囊,也不形成產囊絲,而是以兩個單細胞的營養體、兩個孢子或兩根菌絲的細胞交配直接形成子囊。
低等的子囊菌的子囊是散生的;至於形成產囊絲的較高等的子囊菌,子囊則產生在不同類型的稱作子囊果的孢子果內。根據是否形成子囊果、子囊果的類型和子囊的結構,子囊菌亞門分為六個綱,與植物病害有關的是以下五個綱:
半子囊菌綱(Hemiascomycetes):沒有子囊果。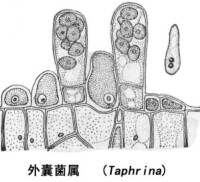
子囊菌亞門真菌
不整囊菌綱(Plectomycetes):子囊果是閉囊殼。
核菌綱(Pyrenomycetes):子囊果是子囊殼。
腔菌綱(Loculoascomycetes):子囊果是子囊座,子囊是雙囊壁的。
盤菌綱(Discomycetes):子囊果是子囊盤。
半子囊菌綱包括許多低等的子囊菌,子囊是散生的,沒有子囊果。它們的營養體是單細胞或不很發達的菌絲體。無性繁殖主要是裂殖或芽殖,不產生真正的分生孢子。有性生殖比較簡單,如前所述,並不形成特殊的配子囊交配,子囊也不是由產囊絲形成的。
(Taphrinales):不形成子囊果,子囊探生於寄主組織表面,無包被,無側絲。
外囊菌目的菌絲體在寄主細胞間過寄生生活,而沒有吸器的形成,其中大多數刺激寄主使呈特殊的變形、膨大、叢生等。它們的主要寄主是羊齒植物、山毛櫸目(Quercifloren)植物和薔薇目(Rosifloren)的植物,但在寄主植物上無子實體形成,它們的子囊直接從營養菌絲中長出來。外囊菌的營養菌絲是雙核的,它們的質配是由兩個子囊孢子或芽孢子來交配的,交配后的孢子萌發即生雙核菌絲。也可能是由單個芽孢子萌發,細胞核在芽管中第一次分裂,形成一對細胞核,以後再發育為雙核菌絲(圖2-25)。這種菌絲體在樹皮上以腐生生活越冬,待到翌春,在樹葉生長之前芽殖細胞在表皮細胞間的胞間層里伸出芽管,菌絲穿入葉片和葉柄組織的細胞間,後來發展到角質層和角質層下生長,形成一層產囊細胞,核配即在該細胞中進行。然後產囊細胞分裂一次,上方1個為子囊母細胞、隨即發育為子囊,下方1個為子囊柄細胞。子囊突破角質層暴露於寄主組織表面,其中有子囊孢子8個。子囊孢子在子囊內或散出后,以芽生方式行無性繁殖,產生大量芽孢子。本目只有l科1屬,即外囊菌科(Taphrinaceae),外囊菌屬(Taphrina)。外囊菌在自然界都是寄生的,子囊平行排列在寄主表面,長圓柱形,子囊孢子圓形、無色。它們通常寄生於木本植物的嫩葉、幼果和芽上,刺激寄主細胞增生,引起過度生長型病變,病葉肥厚皺縮,病果腫大中空,如果芽受害則發育為叢枝。桃縮葉病菌(T. defomans)可為代表。此外,常見的還有李囊果病(T. pruni)、杏、梅縮葉病(T. mume)、櫻桃叢枝病(T. cerasi)及楊樹果腫病(T. somii)等。畸形外囊菌(Taphrina deformans) 畸形外囊菌是以有隔膜的雙核菌絲體在寄主細胞間寄生。有性生殖的過程是雙核菌絲體在寄主角質層下形成一層產囊細胞,其中兩個細胞核進行核配形成一個二倍體的細胞核。產囊細胞伸長突破寄主角質層,細胞核分裂一次,並分隔成兩個細胞,基部的細胞形成子囊柄,上面的細胞形成子囊。因此,畸形外囊菌並不形成特殊的配子囊進行交配,它的子囊是由雙核的菌絲體細胞形成的,子囊平行排列在寄主表面,長圓筒形,其中有8個子囊孢子。子囊孢子單細胞,橢圓形或圓形(圖2-25)。它的無性繁殖是子囊孢子在子囊內外芽殖產生芽孢子,習慣上也將它稱作分生孢子。分生孢子還可以繼續以芽殖的方式繁殖。畸形外囊菌在人工培養基上不形成菌絲體,也是以芽殖的方式繁殖,形成與酵母菌相似的菌落。圖2-25,外囊菌屬 植物葉片表層裸生的子囊 畸形外囊菌以子囊孢子或分生孢子萌發的芽管,從幼葉和幼枝的角質層或氣孔侵入寄主。子囊孢子和分生孢子都是單核的,如何形成雙核的菌絲體還不很清楚。可能是子囊孢子或分生孢子萌發后交配,或分生孢子萌發時細胞核在芽管中分裂為一對細胞核,以後發育成雙核的菌絲體。侵入寄主后,菌絲體先在角質層和表皮下寄生,然後擴展到表皮細胞間和柵欄組織細胞間,刺激葉片柵欄組織和海綿組織的細胞增生和增大,細胞壁變厚,細胞內的葉綠體遭到破壞,葉片表面表現皺縮、肥腫,變為黃褐或紅色等癥狀。葉片表面形成灰白色粉蠟層,即病菌的子囊層。幼枝受害也表現變粗等畸形。
不整囊菌綱真菌的營養體是很發達的菌絲體。子囊散生在閉囊殼的中央。閉囊殼是一種沒有固定孔口的子囊果,它的包被可以是疏鬆的網狀組織到較厚的薄壁組織(圖2-28)。由於子囊壁很早膠化和消解,所以在成熟的子囊果中往往只能看到其中分散的子囊孢子。子囊是產囊絲形成的,一般橢圓形,其中多半有8個圓形的子囊孢子。子囊之間沒有稱作側絲的絲狀體。
許多不整囊菌綱真菌的無性階段很發達,產生大量的分生孢子,在自然界經常看到的是它們的無性階段,如常見的麴黴屬(Aspergillus)和青黴屬(Penicillum)真菌。
散囊菌目
不整囊菌綱只有一個散囊菌目(Eurotiales),有些原來歸在這個綱的真菌如白粉菌,它們的子囊果也是閉囊殼,但子囊是成束著生在閉囊殼的基部,子囊壁也不容易消解,子囊孢子成熟后可從子囊中彈出。因此,目前認為放在核菌綱中更為恰當。
散囊菌目真菌大都是土壤中動植物殘餘組織上的腐生物,其中有些是重要的工業和醫藥微生物,有些引起種子和穀物的霉爛。指狀青黴(Penicillium digitatum)和義大利青黴(P. italicum)是常見的引起柑桔腐爛的重要植物病原物,將在半知菌亞門中介紹。
核菌綱是一類很龐大的真菌,這裡將核菌綱列為四個目,其中與植物病害有關的主要是白粉菌目(Erysiphales)和球殼菌目(Sphaeriales)。
是子囊果有完整的包被和固定孔口的子囊殼,孔口的內側有一層菌絲狀的緣絲。有些核菌綱真菌的子囊果是閉囊殼或近似閉囊殼,有的分類系統將它們歸在不整囊菌綱。至於子囊是雙囊壁的子囊菌,也曾歸在核菌綱中,目前則單獨成為腔菌綱。總的來說,核菌綱應該包括哪些子囊菌以及分為幾個目,意見很不一致。
核菌綱白粉菌目
(Erysiphales):菌絲表生,子囊果無孔口,子囊在閉囊殼中有規則地排列,有些屬只有1個子囊。
白粉菌目的菌絲體大多是表生的,以吸器伸入寄主表皮細胞內吸取養料,很少伸入皮下細胞,大多為外寄生,生活於顯花植物上。有性生殖在寄主表面的鄰近菌絲上生出雄器和產囊器,互相接觸,交配而產生呈球形、扁球形等的閉囊殼,子囊有規則地在閉囊殼內成為一束,或排列成一層,或只有1個子囊。該目中白粉菌科rysiphaceae)最為常見。
白粉菌科菌絲體無色、表生,產生吸器伸入表皮細胞中。有的白粉菌不產生吸器,以它的分枝從氣孔伸入葉肉組織中,它們在葉片的表面形成一薄層菌絲體。無性生殖在分生孢子梗上產生無色、單細胞、橢圓形或長斜方形的分生孢子,孢子單生或串生。菌絲體、孢子梗、粉孢子呈白色或灰色,故當分生孢子大量產生和脫落時,寄主表面就顯出白粉病病徵。分生孢子可以被風吹散,飄落在另一些葉片上,在條件適宜時菌絲髮芽引起新的侵染。這種侵染在夏季可以重複多次。在亞熱帶和熱帶氣候下,往往周年停留在無性階段不斷繁殖。白粉菌的分生孢子可分為三種類型:擬卵孢型(Ovulariopsis)、粉孢型(Oidium)及擬粉孢型(Oidiopsis)(圖2-26)。
圖2-26,白粉病菌分生孢子的三種類型 1,粉孢型 2,擬卵孢型 3,擬粉孢型 ①足細胞 ②孢子母細胞 ③分生孢子
白粉菌的分生孢子,能在濕度很低的情況下萌發,這可能是有些白粉病在很乾燥的環境中仍然猖撅發生的原因之一。在溫帶地區,一般到夏末或秋初,白粉菌開始有性繁殖階段,經雄器與產囊器的配合而產生子囊,子囊在閉囊殼內。閉囊殼球形,閉囊殼的外層細胞變成暗色,其表層的一些細胞發展成為附屬絲。附屬絲有的在閉囊殼中腰環生一圈,也有的生在頂部,它們具有不同的形狀。附屬絲的形狀和子囊的數目是白粉菌分屬的主要依據。子囊果隨病葉在地面越冬,春季吸水破裂,放齣子囊孢子,成為初侵染來源。
中國白粉菌共發現30多個屬,其中6個屬是常見的,它們為害多種園林植物(圖2-27)。
球針殼屬(Phyllactinia)中的桑里白粉病菌(P. corylea),為害桑、梨、柿、核桃等80多種木本植物的葉和新梢。
鉤絲殼屬(Uncinula)中的葡萄白粉病菌(U. necator),為害葡萄的葉、新梢、果梗及果實。槐樹白粉病菌(U. sinensis)為害槐樹葉片,引起粉斑。黃櫨上白粉為害較重,其病原菌是(U. verniciferae)。
單絲殼屬(Sphaerotheca)中的薔薇白粉病菌(S. pannosa),為害薔薇、桃樹的葉片、莖及鳳仙花等,而引起白粉病。
叉絲單囊殼屬(Podosphaera)中的蘋果白粉菌(P. leucotricha),為害蘋果的葉片、幼芽及新梢。此外,日本櫻花和碧桃上也會遭受白粉病的為害,其病原菌為(P. tridaclyla).
根據閉囊殼內子囊的數目及附屬絲的形狀,可對上述6個屬進行檢索。
⒈閉囊殼內產生幾個以致幾十個子囊。
⒉附屬絲柔軟成菌絲狀…………………………… 白粉菌屬(Erysiphe)
⒉附屬絲堅硬,有定型。
⒊附屬絲基部球形,頂端尖銳,呈針狀 …球針殼屬(Phyllactinia)
⒊附屬絲基部不呈球形。
⒋附屬絲頂端二叉狀分枝……………… 叉絲殼屬(Microsphaera)
⒋附屬絲頂端捲曲成鉤狀…………………… 鉤絲殼屬(Uncinula)
⒈閉囊殼內有一個子囊。
⒉附屬絲似菌絲狀,通常不分枝…………… 單絲殼屬(Sphaerotheca)
⒉附屬絲剛直,二叉狀分枝………………叉絲單囊殼屬(Podosphaera)
核菌綱球殼菌目
真菌的子囊果是子囊殼(圖2-28)。子囊殼有球形、半球形或瓶狀的。殼壁(包被)有鮮色肉質、暗色膜質或炭質的。孔口為乳頭狀或長圓柱狀,有緣絲或口須。子囊殼有散生和聚生的,著生在基質的表面或者部分或整個埋在子座內。子囊有圓形、棍棒形、紡錘形或圓柱形,有一層很薄或較厚的囊壁。子囊一般排列成子實層,少數是分散的。子囊之間大都有側絲,但也有很早就消解或沒有側絲的。子囊孢子為單細胞或多細胞,無色或有色,大小差別很大。球殼目真菌一般都有很發達的分生孢子。
圖 2-28,典型的子囊殼的結構
①孔口 ②緣絲 ③子囊殼壁 ④子座 ⑤子囊孔口 ⑥側絲 ⑦子囊 ⑧子囊孢子
無性世代發達,產生各種類型的分生孢子。寄生在植物上的種類,在生長季節多以分生孢子進行繁殖和傳播,到生長後期,或在落葉中產生子囊殼。本目真菌危害植物莖、葉和果實,大多引起壞死性癥狀,也有少數能引起根部腐爛及叢枝、瘤腫等畸形癥狀。下面是幾個能引起園林植物病害的代表性屬。
炭豆菌屬(Rosellinia):子囊殼圓形,有乳頭狀孔口,生於基物表面,有時被一層很薄的子座包圍,炭質或近革質;子囊圓柱形,有側絲;子囊孢子單細胞,橢圓形或梭形,褐色。該菌能為害多種果樹林木,如白紋羽病菌(R.necatrix)是許多闊葉樹根部寄生物,引起根腐。常在樹木病根的表面形成白色菌索和羽毛狀菌絲束。它們以菌絲體及菌核在土壤殘根上越冬,以菌絲穿過土壤傳染(圖2-29)。
圖 2-29,炭豆菌屬 1,子囊殼 2,子囊、子囊孢子及側絲 3,厚垣孢子及其形成 4,分生孢子梗束
小叢殼菌屬(Glomerella):子囊殼叢生在子座上,或半埋於子座內,深褐色,瓶形,殼壁有毛,子囊殼內不形成側絲;子囊棍棒形,無柄,成熟后子囊壁膠化;子囊孢子長圓形,直或略彎,單細胞,無色,萌發時為雙細胞。寄生,引起植物的炭疽病。如七葉樹炭疽病菌(G. cingulata)是最常見的一種,它的寄主很廣,包括蘋果、梨、葡萄、山茶、杉木、女貞等,為害葉、莖及果實。在杉木針葉上較容易產生子囊殼。該屬也能危害梅花的葉片和嫩梢,引起梅花炭疽病(G. mume)(圖2-30)。
圖 2-30,小叢殼屬 子囊殼及子囊
日規殼菌屬(Gnomonia):子座不發達,子囊殼黑色,瓶狀,生於寄主組織中或突破寄主而外露,孔口頸部較長;子囊橢圓形或紡錘形,頂壁厚,中央有溝漕通上孔口,子囊孢子經溝漕射出;子囊孢子雙列,無色,2—4個細胞。常寄生在闊葉樹葉片上引起炭疽病或葉斑病。如懸鈴木炭疽病菌(G. veneta)、榆樹炭疽病(G. ulmea)(圖2-30)。
圖 2-30,日規殼屬埋生在寄主組織中的子囊殼和子囊
腔菌綱分為五個目,其中與植物病害關係較大的是多腔菌目(Myriangiales)、座囊菌目(Dothideales)和格孢腔目(Pleosporales)。
腔菌綱真菌的主要特徵是子囊果是子座性質的。許多單個子囊散生在子座組織中,或者許多子囊成束或成排著生在子座的子囊腔內。這種形式的子囊果稱作子囊座。子囊座形成的過程是產囊絲伸入發育中的子座組織中,隨著子座和子囊的形成,子囊周圍的子座組織消解成子囊腔,子囊就著生在子囊腔內(圖2-35)。因此,子囊腔沒有特殊的腔壁(包被)與子座組織的其它部分分開。子囊座成熟后,子囊腔頂部的細胞組織消解而形成圓形的孔,不象子囊殼那樣很早就形成固定的孔口。同時,子囊之間沒有側絲,有時在子囊之間可看到與側絲相似的絲狀體,這是殘餘的子座組織,一般稱作擬側絲。有的子囊座只有一個子囊腔,子囊腔周圍菌組織的壓縮很象殼壁,表面看起來與球殼目的子囊殼差不多,有人稱它為假囊殼。此外,有些子囊腔是盤狀的。
腔菌綱真菌另一個特徵是子囊是雙囊壁的,外層較薄而沒有彈性,內層則隨著子囊的發育而加厚,特別是頂部更為明顯。內層是有彈性的,頂部往往有缺口或通道。子囊孢子成熟后,外層破裂,內層伸長,子囊孢子就從缺口處或通道彈出。子囊的雙囊壁在子囊孢子釋放時更為明顯。
腔菌綱多腔菌目
該目真菌的特徵是單個子囊散生在子座的薄壁細胞組織中,每個子座中可有許多子囊,並且分佈在不同的層次中。子囊橢圓形。子囊孢子為多格或磚格。這類真菌大都是附生物或寄生物,可以寄生在其它真菌、介殼蟲以及植物的莖葉上。在熱帶和亞熱帶較多,溫帶常見的是多腔菌屬(Myriangium)和痂囊腔菌屬(Elsinoe)。重要的病原物有引起葡萄黑痘病的痂囊腔菌(E. ampelina),引起柑桔瘡痂病的柑桔痂囊腔菌(E. fawcettii)等。
腔菌綱座囊菌目
該目真菌的特徵是成束的子囊著生在有許多子囊腔的墊狀子囊座或很小的子囊殼狀的假囊殼內,子囊之間沒有擬側絲。子囊是倒棍棒狀或短圓筒狀。子囊孢子一般只有一個隔膜。孔口是子座組織溶生的,或者是裂生而有緣絲的。許多座囊菌目真菌是植物葉部的寄生物,特別是在熱帶地區較多。在寄主枯死的黃葉上容易發現它們的有性階段,而在生長期為害的一般是它們的分生孢子階段。其中重要的植物病原物有球座菌屬(Guignardia)、球腔菌屬(Mycosphaerella)、亞球殼屬(Sphaerulina)等,引起葡萄白腐病的葡萄球座菌(G. bidwellii)是其中常見的。
腔菌綱格孢腔目
該目真菌的性狀與座囊菌目相似,所不同的是它們的假囊殼稍大,子囊之間有擬側絲,子囊為圓柱狀,子囊孢子一般是多格的或磚格的(也有單細胞或只有一個隔膜的)。假囊殼一般是單生的,也有聚生的,有的聚生在半埋的子座內,但是很少形成多囊腔的子囊座。格孢腔目真菌有腐生的和寄生的。寄生的大都為害植物的葉片(有的還能為害枝條和果實),一般也是在枯死的枝葉上發現它們的有性階段。生長期為害的是它們的分生孢子階段。黑星菌屬(Venturia)中的蘋果黑星菌(V. inaequalis)和梨黑星菌(V. pyrina)分別引起蘋果和梨的黑腥病,是具有代表性的種。格孢腔菌屬(Pleospora)真菌也是常見的,但它們大都是弱寄生菌。值得提出的還有核腔菌屬(Pyrenophora)和旋孢腔屬(Cochliobolus),它們的分生孢子階段是為害許多禾本科植物的長蠕孢屬(Helminthosporium)真菌。
葡萄座腔菌屬(Botryosphaeria):子座發達,墊狀,黑色,后突破基物外露;子囊腔孔口不顯著,稍有突起;子囊棍棒形,有短柄;子囊孢子8個,單胞,無色,卵圓形至橢圓形(圖2-35)。該病菌大多為害樹木枝幹,果樹枝幹更易受害,在嫁介面處較易得病,引起潰瘍。在晚秋時節,子囊座即在病斑上出現。如蘋果乾腐病菌(B. ribis)能為害蘋果、梨、懸鈴木的主枝和側枝。蘭花葉基腐病菌(B. sp.)為害蘭花葉片,造成基腐。
圖2-35,葡萄座腔菌屬
1,子座組織溶解形成子囊腔 2,子囊 3,成熟的子囊腔內子囊及似側絲
囊孢菌屬(Physalospora):子囊座內有1個成數個子囊腔,子囊腔瓶狀,孔口稍突出;子囊棍棒形或圓柱形;子囊孢子8個,卵圓形或橢圓形,單胞、無色或淡黃色(圖2-36).寄生在枝條、葉片或果實上。如冬青囊孢病菌(P. ilicella)為害冬青葉片.而梨輪紋病菌(P. piricola)主要為害梨、蘋果、杏、木瓜、海棠等植物的枝幹和葉片,果實也能受害。
圖2-36,囊孢菌屬 子囊腔及子囊
球腔圍屬(Mycosphaerella):子囊座中有一個子囊腔,子囊腔黑色,球形或瓶形,有短乳頭狀孔口;子囊棍棒形;子囊孢子橢圓形,無色,雙細胞(圖2-37)。如櫻桃穿孔褐斑病菌(M. cerasella)寄生於櫻桃的葉片上,引起具有同心圈而穿孔性的褐色斑點,直徑1—5mm。鳶尾球腔菌(M. iridis)為害鳶尾葉片。
圖2-37,球腔圍屬
1,子囊腔(示子囊及擬側絲) 2,子囊 3,子囊孢子
煤炱屬(Capnodium):菌絲由圓形細胞聯成串珠狀,常並列結合成束,暗褐色,是植物枝葉表面的腐生物,通常以介殼蟲或蚜蟲分泌的蜜汁為營養來源,由於表生的黑色菌絲體層影響植物光合作用,使植物發生煤污病。常見的有柳煤污病菌(C. salicinum)和柑桔煤污病菌(C. citri)等(圖2-38)。
圖 2-38,煤炱屬
1,葉片受害癥狀 2,病原菌菌絲 3,菌芽 4,長型分生孢子器 5,子囊果, 6,子囊 7,分生孢子器
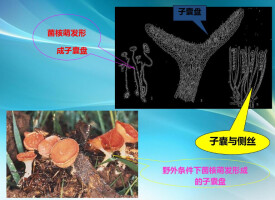
盤菌綱真菌的子囊果是子囊盤,它們的形狀、大小、顏色和質地的差別很大,典型的是盤狀或杯狀,無柄或有柄。子實層是外露的,由排列很整齊的子囊和側絲組成(圖2-40)。有的沒有側絲;有的子囊有囊蓋,子囊孢子從囊蓋釋放;或以唇形開裂代替囊蓋,釋放孢子時,裂口張開;有的子囊沒有囊蓋,子囊孢子從子囊的孔口或裂口釋放。子囊有無囊蓋是區別不同目的性狀之一(圖2-41)。無性繁殖可產生分生孢子,但遠不如其它子囊菌那樣發達。多數不產生分生孢子。盤菌綱真菌大多是腐生的,少數寄生植物。
盤菌綱分為五個目,與植物病害有一定關係的是星裂菌目(Phacidiales)和柔膜菌目(Helotiales)。
盤菌綱星裂菌目
真菌的子囊果開始形成時是一個黑色的子座,圓形、盤形或裂縫形,著生在基質的表面,也有部分或整個埋在寄主組織內的。子實層上有一個由子座組織組成或子座組織與寄主組織結合組成的盾形的蓋,子囊果成熟后,蓋破裂而露齣子實層。子囊果呈子囊盤狀。星裂菌目真菌有腐生和寄生的,其中重要的植物病原物不多,主要的有引起核果葉斑病的冬齒裂菌(Coccomyces biemalis),在中國尚未發現,常見的是槭斑痣盤菌(Rhytisma acerinum)和松針散斑殼(Lophodermiumpinastri)。
盤菌綱柔膜菌目
(Helotiales) 真菌形成比較小或中等大小的有柄或無柄的子囊盤,著生在基質的表面或半埋在基質內,有的是從菌核產生的。子囊棍棒形或圓筒形,子囊無囊蓋,子囊間有側絲。柔膜孢目真菌大都是植物組織上的腐生物,很少發現在土壤或糞堆上,少數是植物的寄生物,如引起多種作物菌核病的核盤菌(Sclerotinia sclerotiorum)和核果褐腐病的核果鏈核盤菌(Monininia laxa)是其中最重要的病原物。有些能引起葉斑,如常見的引起月季黑斑病的薔薇雙殼菌(Diplocarpon rosae)和苜蓿葉斑病的苜蓿假盤菌(Pseudopeziza medicaginis)等。薄盤菌屬(Cenangium):子囊盤自樹皮下生出,無柄或短柄,單質,暗褐色;子囊棍棒狀,內有孢子8個;子囊孢子橢圓形,單細胞,無色;側絲線狀或分叉。危害赤松、黑松、油松,如松枝枯病菌(C. feruginosum)夏秋之間在病死枝條上產生子囊盤,它的無性孢子在中國未曾發現(圖2-31)。
圖 2-31,薄盤菌屬 子囊盤及子囊
念珠盤菌屬(Monilinia):菌絲體與寄主果實形成假菌核,子囊盤自假菌核上產生,漏斗狀或杯狀,有長柄。無性階段很發達,形成叢梗孢屬型分生孢子作為無性世代。在一般條件下,子囊盤很少產生。桃褐腐病菌(M. fructigena)危害桃,受害果實變為僵果,越冬后產生大量分生孢子侵害幼果和嫩梢。該菌也能危害一些薔薇科和杜鵑花科的果實或葉(圖2-32)。
圖 2-32,念珠盤菌屬
1,自僵果上生出的子囊盤 2,子囊及側絲
散斑菌屬(Lophodermium):能引起松樹落針病,它寄生在各種松樹針葉上,是有松落針病菌(L. pinastri)為害的。其分生孢子器和子囊盤都於春季在落葉中形成(圖2-33)。
圖 2-33,散斑菌屬
1,子座及子囊盤 2,子囊及側絲 3,松落針病病葉癥狀
雙殼屬(Diplocarpon):子囊盤在枯死的葉片內形成,有時子囊盤上面具有盾狀子座組織。子囊盤倒三角形,成熟后子實層全暴露。子囊長橢圓形或近乎棍棒形。子囊孢子長形,雙細胞,無色。側絲不分枝。無性階段大多屬於放線孢屬(Actinonema)、盤單隔孢屬(Marssonina)及蟲形孢屬(Entomosporium),寄生葉片,引起多種葉斑病。如薔薇黑斑病菌(Diplocarpon rosae)為害薔薇、月季的葉片,引起黑斑病,全國各地普遍發生。葉片受害產生紫色或黑色、不規則形的病斑,幼枝和花柄也可受害,嚴重時引起落葉(圖2-34)。松落針病和槭漆斑病雖是林木上常見病害,但在中國很少造成較大的損失。