終極腐霉
終極腐霉
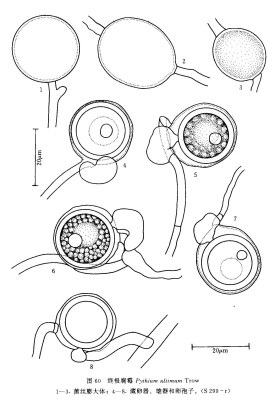
菌落在CMA上呈放射狀。菌絲髮達,分枝繁茂,粗 2.3-9.8μm。菌絲膨大體近球形,多間生,少數切生或頂生,直徑14-32(平均23.4) μm。藏卵器球形、平滑,多頂生,較少間生,罕切生,直徑13-30 (平均22.4)μm。雄器囊狀,彎曲,多同絲生,無柄,緊靠藏卵器形成,偶有下位生和異絲生,7.7-15.5μ m×5.5-10.3μm,平均10.87μm ×6.79μm;授精管明顯可見,粗約1.5μm;每一藏卵器有雄器1(-2)個。卵孢子球形、平滑,不滿器,直徑10-25(平均19.2)μm,壁厚 0.9-2.8(平均1.92)μm,內含貯物球和折光體各一個。
中國分佈:大蔥Allium fistulosum L. :寧夏三一支溝(PM 231)。
韭蔥Allium porrum L. :寧夏中寧(PM 269)。
野油菜Brassica campestris L.:寧夏三一支溝( PM234)。
雪裡蕻Brassica juncea(L.)COSS. var. crispifolia Bail. :寧夏三一支溝(PM 217)。
大白菜Brassica pekinensis Rupr. :寧夏三一支溝(PM229)。
辣椒Capsicum frutescens L. :寧夏中寧(PM274)。
黃瓜Cucumis sativus L.:北京東升(PM 40)。
番茄Lycopersicon esculentum Mill. :寧夏三一支溝(PM 221,252)。
菜豆Phaseolus valgaris L.:寧夏羅家在(PM241, 242)。
蘿蔔Rqphanus sativa L.:寧夏三一支溝(PM247)。
據戴芳瀾(1979)、蔡雲鵬(1991)和戚佩坤( 1994),本菌在我國還為害爪哇白豆寇Amomum compactum Soland ex MATON、泰國白豆寇A. kravanh Pierre ex Gagnep. 、高冠木Calotropis procera(Ait.)Ait. 、木瓜Carica papaya L. 、日本柳杉Cryptomeria japonica D. Don、銀合歡Leucaena leucocephala( Lam.)de Wit. 、楓香樹Liquidambar formosana Hance、球根豆薯Pachyrhizus bulbosus ( L.)Britton、虞美人Papaver rhoeas L.、琉球松Pinus luchuensis Mayr、台灣二葉松P. taiwanensis Hay.、松葉牡丹Portulaca pilosa L. 和蠶豆Vicia faba L.等。
土壤分離:北京海淀(S466,SM303-A);山西大同(S396-C,396-E;江蘇南京(S299-r);貴州貴陽(S420-1);雲南西雙版納(S486);寧夏涇源(SM824),靈武(SM 576).蘆花台 SM 596),中寧(SM 626)等。
世界分佈:土耳其、中國、日本、丹麥、烏克蘭、立陶宛、奈及利亞、加拿大、冰島、吉爾吉斯斯坦、剛果、希臘、拉脫維亞、法國、肯亞、英國、美國、南非、俄羅斯、捷克、斯洛伐克、菲律賓、愛沙尼亞、韓國、紐西蘭、澳大利亞、德國。
Drechsler(1927)說 Pythium ultimum trow常被誤鑒定為P. debaryanum Hesse。Van Luijk(1934)認為Hesse(1874)的P. debaryanum的圖頗為可疑,但他並未推翻其名稱,並認為de Bary(1881)所描述的菌就是P. debaryanum;他寧願用P. debaryanum Hesse em. de Bary這個名稱,而不用P. ultimum。儘管de Bary的描述與 Trow對P. ultimum的描述甚為接近而與Hesse對P. debaryanum的描述則相差甚遠。 de Bary與Trow的描述有一不同點是前者繪圖描述了下位生雄器,而後者卻未提及。但是,Plaats-Niterink(1981)和本文作者認為這種雄器甚為罕見,僅在少數分離物中出現。 P. ultimum與P. paroecandrum Drechsler的區別是前者藏卵器多頂生、缺間生雄器,後者藏卵器多間生並具間生雄器。P. ultimum與其鄰近種的主要區別是它具有袋或囊狀的同絲生雄器、厚壁的卵孢子以及球形的菌絲膨大物和孢子囊。Drechsler(1960a)根據在室溫條件下能釋放遊動孢子而建立了P. ultimum Trow var. sporangiiferum Drechsler變種,而P. ultimum則自動成為原變種P. ultimum Trow var. ultimum。