氨醯
氨醯
氨基酸分子去掉羥基后,剩下的一價原子團統稱為氨醯。當密碼子“GCG”與tRNA上的反密碼子“CGC”配對時,這就意味著tRNA攜帶丙氨酸到核糖體上來了。這不需要經過檢測,因為每種tRNA分子在它到達核糖體之前就與相應的氨基酸結合了。這種結合是在一系列被稱為氨醯-tRNA合成酶作用下完成的。這些酶可以將正確的氨基酸裝載到相應的tRNA分子上,於是,tRNA就可以執行它從DNA的脫氧核糖核苷酸序列信息到蛋白質的氨基酸序列信息的翻譯功能。
大多數細胞可產生二十種不同的氨醯-tRNA合成酶,每一種酶負責一種氨基酸。它們各不相同,各負其責,使一種氨基酸與其相應的一套tRNA分子結合。如圖所示,天冬氨醯-tRNA合成酶(用藍色和綠色表示,兩個tRNA分子以紅色表示)是一個由兩個相同亞基構成的二聚體,其他的酶是或大或小的單體,或是二聚體。有的甚至是一種或多種不同亞基構成的四聚體。某些酶具有相當奇特的構造,例如,絲氨醯-tRNA合成酶(蛋白質編號1SET)。幾乎所有氨醯-tRNA合成酶的結構都可以在蛋白質立體結構資料庫中找到。
正如人們所預料的,這其中的許多酶識別tRNA分子的反密碼子。但在某些情況下有所不同。以絲氨酸為例,它有六種不同的密碼子,所以絲氨醯-tRNA合成酶必須能識別帶有六種不同反密碼子的tRNA分子,包括完全不同的“AGA”、“GCU”。於是,酶不得不辨別tRNA分子上的氨基酸接受臂和其他部位的鹼基,尤其是在tRNA序列編號為73的鹼基,似乎在很多情況下起著極其重要的作用,因此被稱為區別鹼基。但有時,它可以完全被忽略。應注意的一點是,每個酶在正確識別tRNA分子的同時,必須阻止錯誤組合。因此,每一個tRNA分子都有一套促進與正確的酶結合的積極因子,也有一套抑制不適當結合的消極因子。例如,天冬氨醯-tRNA合成酶,如圖所示(蛋白質編號1ASZ)能識別tRNA分子的區別鹼基和反密碼子周圍的四個鹼基,而另一個鹼基,鳥嘌呤37,不用於識別,但必須通過甲基化保證tRNA分子不會與精氨醯-tRNA合成酶錯誤結合。
近來,對整個基因組分析的結果,讓人們非常吃驚:有些有機體並不存在全部編碼這二十種氨醯-tRNA合成酶的基因。但它們確實是用這二十種氨基酸去構建蛋白質的。有機體對這個矛盾的解決表明,有更複雜的機制存在。這種現象經常出現。例如,某些細菌不存在將谷氨醯胺裝載到相應tRNA上的合成酶。但它可以用一種酶將谷氨酸加到所有的谷氨酸tRNA分子上和所有的谷氨醯胺tRNA分子上,然後,另一種酶作用於谷氨醯胺tRNA分子,將谷氨酸轉變為谷氨醯胺,從而形成正確的配對組合。
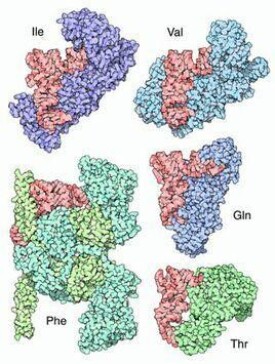
了五種氨醯-tRNA合成酶與其tRNA分子的複合體。tRNA分子(以紅色表示)以相同的方向排列。注意,不同的酶是從不同的角度接近tRNA分子的。作用於異亮氨酸(蛋白質編號1FFY)、纈氨酸(蛋白質編號1GAX)和谷氨醯胺(蛋白質編號1EUQ)的合成酶像是tRNA分子的搖籃,抓住反密碼子環(在tRNA分子底部),並將氨基酸接受臂(在tRNA分子右部頂端)置於它的活性部位。這些酶有相似的蛋白質結構,類似的作用方式,並且都將氨基酸加在tRNA分子末端核糖的2'-羥基上,被稱為“類型Ⅰ”酶。而作用與苯丙氨酸(蛋白質編號1EIY)和蘇氨酸(蛋白質編號1QF6)的酶屬於“類型Ⅱ”,它們從另一側接近tRNA,並將氨基酸加到tRNA末端核糖的3'-羥基上。
氨醯-tRNA合成酶執行其功能時,必須有很高的準確性。它們如果犯了錯誤,將會導致氨基酸在蛋白質合成中的錯誤定位。這些的出錯率為萬分之一。對於絕大多數氨基酸,這個準確率不難達到。因為,各個氨基酸之間有很大的不同。而且,前面曾提到過,不同tRNA分子的許多部位都可用於正確識別。但是,在某些情況下,酶難於選擇正確的氨基酸而必須求助於特殊機制。
異亮氨酸正是這樣一個例子。在酶中,有一個能識別異亮氨酸的小洞,這個洞太小,而不宜讓像甲硫氨酸和苯丙氨酸這樣的大分子進入,並且,這個洞是疏水的,帶有極性側鏈的氨基酸,也被阻擋在外。但是比異亮氨酸稍小的纈氨酸,由於僅比它少了一個甲基,剛好能進入這個口袋。纈氨酸代替異亮氨酸的幾率為一百五十分之一。這個錯誤率太高了。於是必須有校正機制出馬。異亮氨酸合成酶(蛋白質編號1FFY)用有校正作用的另一個活性部位來解決這個難題。錯誤的纈氨酸,而不是異亮氨酸能靠近這個活性部位。這樣,纈氨酸被水解掉,留下tRNA分子,等待正確的異亮氨酸。這個校正步驟將總的錯誤率降到三千分之一。
基本信息
- 中文名
- 氨醯
- 所屬學科
- 化學學科
- 定義
- 一價原子團
- 酶
- 20餘種
- 性質
- 準確性