染色體變異說
染色體變異
在真核生物的體內,染色體是遺傳物質DNA的載體。當染色體的數目發生改變時(缺少,增多)或者染色體的結構發生改變時,遺傳信息就隨之改變,帶來的就是生物體的後代性狀的改變,這就是染色體變異。它是可遺傳變異的一種。根據產生變異的原因,它可以分為結構變異和數量變異兩大類。
染色體變異會導致遺傳病,常見病例21-三體綜合征等,極少數為有利變異,比如三倍體植株培育等。
染色體結構變異最早是在果蠅中發現的。遺傳學家在1917年發現染色體缺失,1919年發現染色體重複,1923年發現染色體易位,1926年發現染色體倒位。人們在果蠅幼蟲唾腺染色體上,對各種染色體結構變異進行了詳細的遺傳學研究。
光學顯微鏡下可見染色體結構的變異或者染色體數目變異,染色體結構變異的發生是內因和外因共同作用的結果,外因有各種射線、化學藥劑、溫度的劇變等,內因有生物體內代謝過程的失調、衰老等。在這些因素的作用下,染色體可能發生斷裂,斷裂端具有癒合與重接的能力。當染色體在不同區段發生斷裂后,在同一條染色體內或不同的染色體之間以不同的方式重接時,就會導致各種結構變異的出現。下面分別介紹這幾種結構變異的情況。
⑴最初細胞質存在著無著絲點的斷片。
⑵缺失雜合體中,聯會時形成環狀或瘤狀突起,但易與重複相混淆,必須:
①參照染色體的正常長度
②染色粒和染色節的正常分佈
③著絲點的正常位置
⑶當頂端缺失較長時,可在雙線期檢查缺失雜合體雜交尚未完全端化的二價體,注意非姐妹染色單體的末端是否有長短。
遺傳效應:缺失對個體的生長和發育不利:
①缺失純合體很難存活
②缺失雜合體的生活力很低
③含缺失染色體的染色體一般敗育
④缺失染色體主要是通過雌配子傳遞的
缺失是指染色體上某一區段及其帶有的基因一起丟失,從而引起變異的現象。缺失的斷片如系染色體臂的外端區段,則稱頂端缺失;如系染色體臂的中間區段,則稱中間缺失。缺失的純合體可能引起致死或表型異常。在雜合體中如攜有顯性等位基因的染色體區段缺失,則隱性等位基因得以實現其表型效應,出現所謂假顯性。在缺失雜合體中,由於部分缺失引起的遺傳效應隨著缺失片段大小和細胞所處發育時期的不同而不同。在個體發育中,缺失發生得越早,影響越大缺失的片段越大,對個體的影響也越嚴重,重則引起個體死亡,輕則影響個體的生活力。在人類遺傳中,染色體缺失常會引起較嚴重的遺傳性疾病,如貓叫綜合症等。缺失可用以進行基因定位。
染色體上增加了相同的某個區段而引起變異的現象,叫做重複。分為順接重複和反接重複。前者指的是某區段按照染色體上的正常順序重複,後者指的是重複時顛倒了某區段在染色體上的正常順序。在重複雜合體中,當同源染色體聯會時,發生重複的染色體的重複區段形成一個拱形結構,或者比正常染色體多出一段。重複引起的遺傳效應比缺失的小。但是如果重複的部分太大,也會影響個體的生活力,甚至引起個體死亡。例如,果蠅的棒眼就是X染色體特定區段重複的結果。重複對生物體的不利影響一般小於缺失,因此在自然群體中較易保存。重複對生物的進化有重要作用。這是因為“多餘的基因可能向多個方向突變,而不致於損害細胞和個體的正常機能。突變的最終結果,有可能使“多餘的基因成為一個能執行新功能的新基因,從而為生物適應新環境提供了機會。因此,在遺傳學上往往把重複看做是新基因的一個重要來源。
鑒別:⑴重複區段較長時:在雜合體中重複區段的二價體會突出環或瘤,不能同缺失雜合體的環或瘤相混淆。
⑵重複區段很短時:可能不會有瘤或環出現。果蠅唾腺染色體體細胞聯會時特別大,其中出現很多橫紋帶,可以作為鑒別缺失和重複的標誌。
遺傳效應:1、擾亂基因的固有平衡體系
2、重複引起表現型變異:
⑴基因的劑量效應:細胞內某基因出現次數越多,表現型效應越顯著
⑵基因的位置效應:基因的表現型應因其所在的染色體不同位置而有一定程度的改變
指某染色體的內部區段發生180°的倒轉,而使該區段的原來基因順序發生顛倒的現象。倒位區段只涉及染色體的一個臂,稱為臂內倒位;涉及包括著絲粒在內的兩個臂,稱為臂間倒位。倒位的遺傳效應首先是改變了倒位區段內外基因的連鎖關係,還可使基因的正常表達因位置改變而有所變化。倒位雜合體聯會時可形成特徵性的倒位環,引起部分不育性,並降低連鎖基因的重組率。倒位雜合體形成的配子大多是異常的,從而影響了個體的育性。倒位純合體通常也不能和原種個體間進行有性生殖,但是這樣形成的生殖隔離,為新物種的進化提供了有利條件。例如,普通果蠅的第3號染色體上有三個基因按猩紅眼—桃色眼—三角翅脈的順序排列(St—P—Dl);同是這三個基因,在另一種果蠅中的順序是St—Dl—P,僅僅這一倒位的差異便構成了兩個物種之間的差別。
鑒別:⑴較短倒位區段:倒位雜合體聯會的二價體在倒位區段內形成“倒位圈”
⑵較長倒位區段:倒位區反轉過來與正常染色體的同源區進行聯會,倒位區段以外的部分只有保持分離。
遺傳效應:1、倒位雜合體的部分不育
2、位置效應:倒位區段內、外基因之間的物理距離發生改變,其遺傳距離一般也改變。
3、降低倒位雜合體上連鎖基因的重組率
4、倒位可以形成新種,促進生物進化
易位是指一條染色體的某一片段移接到另一條非同源染色體上,從而引起變異的現象。如果兩條非同源染色體之間相互交換片段,叫做相互易位,這種易位比較常見。相互易位的遺傳效應主要是產生部分異常的配子,使配子的育性降低或產生有遺傳病的後代。易位雜合體在減數分裂偶線期和粗線期,可出現典型的十字形構型,終變期或中期時則發展為環形、鏈形或∞字形的構型。易位的直接後果是使原有的連鎖群改變。易位雜合體所產生的部分配子含有重複或缺失的染色體,從而導致部分不育或半不育。例如,慢性粒細胞白血病,就是由人的第22號染色體和第14號染色體易位造成的。易位在生物進化中具有重要作用。例如,在17個科的29個屬的種子植物中,都有易位產生的變異類型,直果曼陀羅的近100個變種,就是不同染色體易位的結果。
遺傳效應:1、半不育是易位雜合體的突出特點
①相鄰式分離:產生重複、缺失染色體,配子不育
②交替式分離:染色體具有全部基因,配子可育
2、降低臨近易位接合點的基因間重組率
3、易位可以改變基因連鎖群
4、造成染色體融合而改變染色體數目
染色體結構變異,使排列在染色體上的基因的數量和排列順序發生改變,從而導致性狀的變異,大多數染色體變異對生物體是不利的,有的甚至導致死亡。
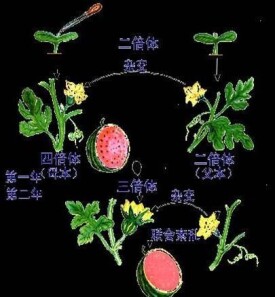
指以一定染色體數為一套的染色體組呈整倍增減的變異。一倍體只有1個染色體組,一般以X表示。二倍體具有2個染色體組。具有3個或3個以上染色體組者統稱多倍體,如三倍體、四倍體、五倍體、六倍體等。單倍體是體細胞中染色體數為本物種配子染色體數目的個體。一般奇數多倍體由於減數分裂不正常而導致嚴重不孕。如果增加的染色體組來自同一物種,則稱同源多倍體。如直接使某二倍體物種的染色體數加倍,所產生的四倍體就是同源四倍體。如使不同種、屬間雜種的染色體數加倍,則所形成的多倍體稱為異源多倍體。異源多倍體系列在植物中相當普遍,據統計約有30~35%的被子植物存在多倍體系列,而禾本科植物中的異源多倍體則高達75%。栽培植物中有許多是天然的異源多倍體,如普通小麥為異源六倍體、陸地棉和普通煙草為異源四倍體。多倍體亦可人工誘發,秋水仙素處理就是誘發多倍體的最有效措施。
生物體的2n染色體數增或減一個以至幾個染色體或染色體臂的現象。出現這種現象的生物體稱非整倍體。其中涉及完整染色體的非整倍體稱初級非整倍體;涉及染色體臂的非整倍體稱次級非整倍體。在初級非整倍體中,丟失1對同源染色體的生物體,稱為缺體(2n-2);丟失同源染色體對中1條染色體的生物體稱為單體(2n-1);增加同源染色體對中1條染色體的生物體稱為三體(2n+1);增加1對同源染色體的生物體稱為四體(2n+2)。在次級非整倍體中,丟失了1個臂的染色體稱為端體。某生物體如果有1對同源染色體均系端體者稱為雙端體,如果1對同源染色體中只有1條為端體者稱為單端體。某染色體丟失了1個臂,另1個臂複製為2個同源臂的染色體,稱為等臂染色體。具有該等臂染色體的生物體,稱為等臂體。等臂體亦有單等臂體與雙等臂體之分。由於任何物種的體細胞均有n對染色體,因此各物種都可能有n個不同的缺體、單體、三體和四體,以及2n個不同的端體和等臂體。例如普通小麥的n=21,因此它的缺體、單體、三體和四體各有21種,而端體和等臂體則可能有42種。染色體數的非整倍性變異可引起生物體的遺傳不平衡和減數分裂異常,從而造成活力與育性的下降。但生物體對染色體增加的忍受能力一般要大於對染色體丟失的忍受能力。因1條染色體的增減所造成的不良影響一般也小於1條以上染色體的增減。非整倍性系列對進行基因的染色體定位、確定親緣染色體組各成員間的部分同源關係等,均具有理論意義。此外,利用非整倍體系列向栽培植物導入有益的外源染色體和基因亦有重要的應用價值。如小麥品種小偃759就是普通小麥增加了1對長穗偃麥草染色體的異附加系,而蘭粒小麥則為普通小麥染色體4D被長穗偃麥草染色體4E所代換的異代換系。
許多癌症如肺癌、腸癌等都是由於其腫瘤細胞中染色體數目變異所造成的,而且科學家們也發現細胞調控因子或者紡錘體蛋白的突變會造成染色體不分離(chromosome nondisjunction,即細胞分裂進入中後期時,某一對同源染色體或者姐妹單體未分別移向兩極,造成子細胞中一個染色體數目增多,一個減少的現象),從而引起染色體數目變化。
在繁殖過程中減數分裂的時候同源染色體沒有分開,或者有部分沒有分開,也會導致生殖細胞中染色體對數發生改變。
當前,在染色體、基因與犯罪關係的研究中,性染色體異常理論引起了人們的較大關注。這種理論認為,性染色體為XYY型的染色體異常與暴力犯罪有關。人類的體細胞中共有23對染色體,人的所有遺傳信息和密碼都貯藏在這23對染色體中。其中,前22對稱為“常染色體”,其在男女兩性中都是相同的。第23對染色體稱為“性染色體”,具有兩種形式,其中女性“性染色體”的組成為XX,男性為XY。但是如果初級卵母或精母細胞在減數分裂時發生錯誤,性染色體不分離,就會產生有兩條性染色體的卵子或精子,由這樣的卵子或精子發育成的個體染色體總數為47條,有三條性染色體,這在醫學上叫做47XYY綜合征,患該症的病人是1961年在英國首次被發現的,該病的發生率約為1:1000。
這種多一條Y染色體的XYY型男性被稱為“超男性”,他具有身材高大、肌肉發達、動作不協調、自控能力差、脾氣暴烈等生理特徵,易衝動和實施攻擊性暴力行為,尤其是在青春期。1968年芝加哥一個名叫斯貝克的罪犯連續殺死了8名護士,后經分析研究,發現該罪犯具有XYY異常性染色體。近年來曾引起轟動的聶樹斌強姦殺人案的真兇王書金,就被懷疑為暴力性犯罪染色體異常者,他多次實施強姦犯罪並殺死了其中的4人,最早的一起是他15歲時在自己村裡強姦了一個來走親戚的小女孩並被判處有期徒刑3年。另外,就整體而言,XYY型男性並不一定存在智力障礙問題,但是如伴有智能缺乏和中樞神經系統異常,則更容易產生犯罪的傾向,容易進行殺人和性犯罪,攻擊性極強,且不能自制。
1965年,英國的傑可布斯等人研究了197個犯罪人,發現其中很大一部分人的染色體屬於XYY的異常態,因此就認為,“超男性”染色體中多出的一個男性染色體在某種程度上與這類男性的暴力行為有關,是導致他們實施犯罪的生理因素。根據相關統計數字,XYY型男性中刑事犯罪率較正常人群為高,大約可達幾到十幾倍,在普通群體中,每1000個男性中具有兩條Y染色體的僅有1人,而在一些監獄的犯人中,這個比率高出了5倍,在極個別監獄中這個比率甚至會高出近20倍。但是迄今為止有關研究尚不能解釋說明,為什麼並非所有暴力犯罪人的性染色體都是“超男性”,而大多數性染色體異常的人並未實施犯罪,以及女性暴力犯罪現象的存在。
基本信息
- 中文名
- 染色體變異
- 外文名
- chromosomal variation
- 原因
- 染色體缺失,增添,易位或倒位
- 常見病例
- 21-三體綜合征,貓叫綜合征等
- 產生後果
- 遺傳病,極少數為有利變異
- 發生時期
- 有性生殖形成配子時
- 生物類別
- 真核生物
- 有利應用
- 三倍體植株(如無籽西瓜)培育