物種形成
物種形成
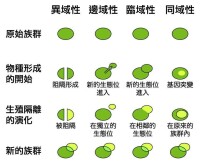
物種形成,又稱為種化,是演化的一個過程,指生物分類上的物種誕生。自然界的物種形成主要有4種模型——異域性物種形成(Allopatric speciation)、同域性物種形成(Sympatric speciation)、邊域性物種形成(Peripatric speciation)與臨域性物種形成(Parapatric speciation)。
物種形成(speciation)是新物種從舊物種中分化出來的過程,即從一個種內產生出另一個新種的過程,包括三個環節:突變以及基因重組為進化提供原料,自然選擇是進化的主導因素,隔離是物種形成的必要條件。
物種形成
同域種化:同一物種在相同的環境,由於行為改變或基因突變等原因而演化為不同的物種。
邊域種化:種化過程中,一個小族群由於某種原因和原來的大族群隔離;隔離時,小族群的基因經歷劇烈變化;當小族群再跟大族群相遇時,已經形成不同物種。
鄰域種化:兩個種化中的族群雖然分開,但是相鄰;從一極端到另一極端之間的各族群都有些許不同,但彼此相鄰的兩族群之間仍能互相雜交;不過,在兩邊最極端的族群已經差異太大而形成不同的種類。
現代生物學關於種形成的研究對象大都是進行有性生殖的動植物,因而種形成的研究多集中於“生殖隔離”的起源問題上,即研究在一個種的群體內如何產生或分化出與原群體生殖上隔離的亞群體,後者就是一個新種的開始。
C·R·達爾文早就注意到地理因素在種分歧和種形成中的作用。P·M·哈蒙德1981年將與種形成有關的地理因素區分為13種情況,並歸納為4種模式:分佈區重疊模式、分佈不重疊模式、分佈區相鄰模式、分佈區不重疊—相鄰模式。
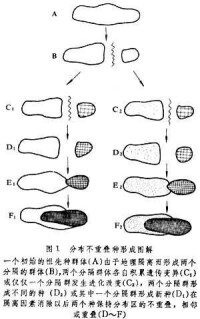
所謂“分佈區重疊”是指形成不同種的原群體在地理分佈上是連續的,所謂“分佈不重疊”是指形成不同的種的原群體由於某種地理隔離因素而被分隔,所謂“分佈區相鄰”是指形成不同種的原群體之間有不完全的地理隔離。區分種形成方式主要依據群體初始分化和生殖隔離完成過程中的地理分佈情況,至於生殖隔離完成(種形成過程結束)以後地理分佈情況則並不重要。一般說來可以區分為三種不同的種形成方式:①分佈區不重疊的種形成,②分佈區重疊的種形成,③分佈區相鄰的種形成。
此外,根據種形成過程的特點還可區分出兩種不同的種形成方式:①漸變的種形成,②量子種形成。分佈區不重疊和相鄰的種形成過程一般是漸變的,分佈區重疊的種形成過程往往是“跳躍”的,即“量子種形成”方式。
分佈區不重疊的種形成
物種形成
不重疊種形成過程通常是長時間的,不可能自始至終直接觀察到。這種物種形成方式的提出,多半是根據間接的證據,例如不完全種(族、亞種或半種)的存在,某些生物種的地理分佈等。一些大洋中的群島對於某些陸地生物來說是不重疊種形成的合適環境。最典型的例子是達爾文曾考察過的南美西岸的加拉帕戈斯群島上的種類豐富而又歧異的動植物區系。例如被稱為達爾文鶯的小鳥有10~14種之多,達爾文認為這些不同種的鶯來自一個從大陸遷徙到該群島的祖先種。由於群島與大陸隔離和群島之間的隔離使得遷入的鳥群分異,並形成不同的種,加拉帕戈斯群島上有3種龜,分別棲居於被海洋分隔的3個小島上,這也是地理隔離因素所促成的不重疊種形成過程的間接證據。
在海洋生物中也有許多由於地理隔離而形成不同亞種的實例。某些物種在北太平洋東西兩側有不連續的分佈,例如某些軟體動物和魚類在北美西海岸是一個亞種而在蘇聯遠東地區和日本北部沿海是同種的另一個亞種,原因是從白令海峽流過的來自北極的寒冷海流,構成了海洋生物的分佈障礙。某些海洋動物在北大西洋和北太平洋對稱地分佈而在北極中斷。例如鱈魚分佈於北大西洋,在北太平洋則有同種的另一亞種,但在北極卻沒有鱈魚。顯然北極的寒冷是分佈中斷的原因,但可能在某一地質時期北極地區曾經是比較溫暖的,兩大洋北部的生物曾有連續的分佈。中美的某些生物,由於巴拿馬地峽的阻隔,而沿著東西兩岸分佈著兩兩對稱的姐妹種。所謂姐妹種就是形態上非常相似而生殖上隔離的完全種。在地峽形成以前它們的分佈可能是連續的。
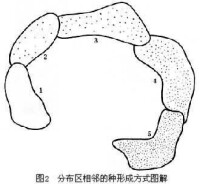
分佈區相鄰的種形成
物種形成
由若干相鄰分佈的亞種組成的廣布種,各相鄰的亞種之間有一定的基因交流,但分佈兩端的亞種(1與5)通過長期的遺傳差異的積累而可能達到生殖隔離(即使分佈區重疊也不能雜交)。
分佈區重疊的種形成
一些證據表明,生殖隔離可以由於生物種的行為差異、發情或開花季節差異、寄主,以及其他原因而導致在那些分佈區並非完全不重疊的群體內產生。有些學者提出一些證據證明植物的某些新種的形成是沒有地理隔離的情況下達到生殖隔離的。
但分佈區是否重疊往往很難區分。例如人體寄生有3種虱:陰虱寄生於腋毛和陰毛中,人虱分佈於軀體(主要在衣服上),頭虱生於頭髮上。若從整個人體看,3種虱的分佈區是重疊的,但若從人體的局部部位看,它們又是不重疊的。分佈區是否重疊一般應根據具體的生物對象的活動能力和行為等特點來判斷。
分佈區不重疊的和相鄰的種形成方式
新種產生於遺傳上連續的單個群體之內(分佈區重疊的種形成)或者產生於有基因交流的相鄰分佈的一系列群體之中(分佈區相鄰的種形成)。在沒有任何阻止基因交流的外在的隔離因素的情況下,新種從母群體中分化產生出來的一些證據多是從觀察研究“成對種”和“初始種”的分佈和區分特徵而獲得的。“成對種”和“初始種”被認為是新形成的或形成中的物種,它們常是研究物種形成的對象。但是根據它們的地理分佈和區分特徵來推斷種形成方式還存在許多問題。例如“成對種”或“初始種”的現有的分佈區相鄰的狀況也可能是種間競爭所造成的地理分佈上的相互排斥,如是,則其相鄰的分佈乃是種形成的結果或種形成后的現象。母種與新種之間的區分特徵中哪些是種形成過程所涉及的,哪些是種形成以後產生的,也很難判斷。
根據學者們提供的資料,可將分佈區重疊的和相鄰的種形成歸納為兩種可能的方式:
①兩階段的種形成過程。在一個遺傳上連續的群體之內首先發生種內分異,經過相當長時間的種內分異的積累而最終在群體內產生生殖隔離的新種。
②一步“跳躍”的過程。新種從母群體中突然產生,即通過染色體突變或基因突變或其它機制在較短時間內產生與母群基因交流阻斷的新類型或新種,一舉而達到生殖隔離。
關於第一種方式可以舉出下面一些事實。分佈區重疊的種形成的實例如某些寄主專一的昆蟲寄主族是近期形成的半種,是種內分異的結果。
1975年,G·L·布希在研究果蠅的寄主族(指同一物種的有不同的寄主專一性的族群)后指出:在同一物種內寄主的轉換可以造成分佈區重疊的寄主族。布希認為,對於果蠅來說寄主轉換隻需要很少的基因突變,而寄主的改變只是種內分異的第一步,其後是生態小生境的改變並伴隨寄主族之間、新寄主族與母群體之間的生殖的障礙。例如,果蠅的交配只發生在其寄主的果實上,不同的寄主族之間發生交配的可能性很小,這樣,寄主族之間的分異就會和地理隔離造成的效果一樣。
對澳洲蝗蟲的研究提供了分佈區相鄰的種形成方式的例證。1978年,M·J·D·懷特研究了澳洲蝗蟲的相鄰分佈的近緣種和染色體族之後提出了“原地靜態”的種形成方式,他認為,染色體重排(染色體融合、分離、倒位、易位、斷裂、缺失等)在種內初始分異中起重要作用。澳洲蝗蟲是失去飛翔能力的、種內核型變異很大的昆蟲,大約有200種之多。在一些種(例如在Vandiemenella viatica)之內有許多核型不同的、地理上相鄰分佈的染色體族。有一些核型不同的近緣種的分佈區也是相鄰的。這些相鄰分佈的族或種的分佈區往往有明確的邊界,互不重疊。研究還發現染色體族之間存在著程度不同的生殖障礙。例如,各族之間的雜合子不育或生育力低,從而在各染色體族之間形成一狹窄的“雜交帶”。由於基因交流的阻隔,染色體族有可能發展為生殖隔離的新種。
但對於這些分佈區相鄰的染色體族和種的形成的解釋還有一些不同的看法。首先是關於染色體重排在種形成過程中究竟起多大作用還難以評價。在澳洲蝗蟲中並非所有的近緣種在核型上都有區別。另一個疑問是:相鄰的分佈是否可能為種間競爭排斥的結果,即種形成后的現象。還有一個問題是:種內分異所造成的族(半種)之間的生殖障礙(不完全的生殖隔離)如何發展為完全的生殖隔離。達爾文和A.R.華萊士都曾提出過生殖障礙可以通過自然選擇而逐步發展的假說。如果種內分異產生出多種表型,如果中間類型(雜種)的適應值相對較低,那麼通過“中間類型”或“雜種”的淘汰絕滅而由生殖障礙發展為完全的生殖隔離。
第二種跳躍的種形成方式是不經過種內分異而一步達到生殖隔離的過程。例如在植物中,多倍體的種可能是從其二倍體的親種中快速產生的。H.劉易斯(1966年)認為木本植物的染色體基數在科或亞科級的大分類單元中是恆定的,在草本植物中染色體基數在屬一級的類群中是恆定的。植物的染色體基數的改變可能涉及高級分類群的形成,因而是跳躍的。

物種形成
古生物學家G·G·辛普森(1944年)鑒於化石記錄中某些高級分類單元在較短的地質時間內出現而提出“量子進化”的概念。他認為生物種通過跳躍式的“量子進化”就能越過適應閾限,進入新的適應帶,從而發生適應輻射和產生高級分類單元。但辛普森認為“量子進化”可以通過分枝的種形成,也可以通過線系進化而實現。
某些生物學家(例如G·G·辛普森、E·邁爾等)認為,除了漸變的種形成方式以外,還可能存在快速的、跳躍式的種形成方式。另一些生物學家(如斯坦利)則認為種形成不可能是漸變的過程,只能是“跳躍”式的、“相對快速”的過程。處於“相對穩定”狀態的種,通過“突然”的改變而分化出新種的過程稱為量子形成。
1963年,這個詞最先由V·格蘭特提出,S·M·斯坦利把它定義為“大部分進化集中發生在種形成的初期的相當短的時間(相對於種的壽命而言)之內的種形成過程”。關於種形成的方式是漸變論者與間斷平衡論者之間爭論的焦點。漸變論者認為漸變的種形成是一般的、普通的,快速、跳躍式的種形成是極特殊的情況;間斷平衡論者則認為種的形成只能是“跳躍”式的。
目前,關於量子種形成問題的爭論焦點在於快速種形成的具體機制。一些學者強調這種機制是“大突變”,甚至認為一個或少數幾個突變就能產生一個新種或新的高級分類單元。這實際上又回到了20世紀初期德·弗里斯的“突變論”。
有的學者則提出另一種快速種形成的可能機制:在分隔狀態下的小群體,隨機因素有助於快速進化和種形成。如果一個廣布的種由許多小群體組成,小群體之間有不完全的隔離,則隨機因素會造成小群體對總體的大的偏離和小群體之間的遺傳差異。但由於小群體的不穩定也增大了絕滅的可能性。
神學家或信奉神學的科學家認為一切物種都是由上帝創造的。譬如,十八世紀的分類學家林奈起初認為萬能的神創造了所有的物種。到了晚年,林奈的思想有所變化,認為上帝創造了植物的目(order),在目以下的階元一直到種所有的分類單位都是由雜交(“混合”)形成的(邁爾1990)。
法國人莫培兌(Pierre Louis Moreau de Maupertuis,1698-1759)雖以數學和天文學為專業,但對生物現象卻很感興趣。他認為自然界的多樣性和異質性是如此之多,因此世界不可能是按設計產生的,而傾向於盧克萊修和伊壁鳩魯學派所提出的起源主要出自偶然的觀點。他曾說,“我們能不能這樣來說明怎樣只從兩個個體形成了形形色色的各種物種?它們的最初來源很可能只是偶然的產物,其中基本的微粒不像在父本和母本親代中那樣有次序;每一種程度的差誤就可能產生一個新種。通過反覆多次的偏差就可能形成我們現在所見到的生物無窮無盡的多樣性”(邁爾1990)。
博物學家拉馬克認為每一進化路線是由簡單生物單獨的自然發生形成的,簡單生物隨後就進化成高等生物。博物學家達爾文相信,“一切動物和植物都是從某一種原始類型傳下來的”,關於物種分化的內在機制他並未深入涉及,他曾說,“自然選擇就經常傾向於保存任何一個物種的最分歧的後代”(Darwin 1872),但並未就這一論述給出任何道理。
哲學家柏格森(1907)曾精闢地說,“生命從起源開始,就是同一個衝動的延續,它分成了各種不同的進化路線。通過一系列的增添,一些東西成長發展起來,而這些增添就是許許多多的創造。這種發展造成了各種趨勢的分離,這些趨向若是不變得互不相容,其成長就不能超過某個點……進化實際上出現在數百萬的個體上,發生在各個不同的路線上,每條線路都結束於一個岔口,而從這個岔口又發射出一些新的路徑,如此發展,乃至無窮”。雖然柏格森對進化的可能場景描繪得相當精彩,但這個“衝動”到底是什麼呢?
進化遺傳學家杜布贊斯基(1972)說,“對生物如此豐富的多樣性,有沒有一個合理的解釋呢?……許許多多生物界的古怪玩意,是從哪裡來的呢?唯一合理的解釋,就是有機體多樣性是對應於地球上環境的多樣性而進化來的。沒有一個物種,不管是多麼完美和多能,能夠開發所有的生存機會……化石證據清清楚楚地顯示了,大多數進化途徑逐漸走向滅絕。現存的有機體只是過去生存過的少數物種的成功後代-如果你找得越遠,這個比例會越來越小。然而,存活物種的數目並沒有縮減,事實上,它可能隨著時間的推移而越來越多。所有這一切,只有根據進化論才能夠明白。如果是上帝從虛無之中造出了無數物種,然後讓其中的大多數滅絕,這是多麼沒有道理的操作”!
謝平(2014)提出了物種形成的綜合機制,認為物種是遺傳學、生理學和生態學機制綜合作用的產物。
遺傳學機制—有性生殖物種基因庫的反覆分裂
新物種的創造肯定不是舊物種的有意識的或目的性的行為。從本質上來看,地球上如此巨大的物種創造現象似乎只是有性生殖方式的“意外”之物。任一物種的存在與演化都是通過生殖來實現的,自然界的生殖方式雖然千差萬別,但都可以歸結為兩種—無性生殖和有性生殖,而這兩種生殖方式又是由兩種細胞分裂方式—減數分裂和有絲分裂(原核生物的細胞分裂方式比有絲分裂更為簡單)為平台來操作的。地球上98%以上的物種中能進行有性生殖,雖然所有的原核生物只能進行無性生殖,以及一些植物和低等的無脊椎動物能進行有性與無性的混合生殖方式,但能進行無性生殖的生物種類遠不及有性生殖。顯然,有性生殖與無性生殖方式在物種中的分佈存在巨大的差別,因此,有理由判斷,有性生殖是推動物種分化的重要生殖方式。這是為什麼呢?這取決於以下的有性生殖的生態遺傳學本質。
首先,有性生殖物種是將基因庫(承載著種族延續的遺傳指令)流動性地保存於相互間能夠交配的種群的個體之中。任何一個個體都是來自基因庫中的兩個個體(父本和母本)的基因組的隨機組合。
第二,與無性生殖相比,有性生殖物種必定具有產生豐富變異的遺傳機制。現已知道,有性生殖物種在減數分裂過程中同源染色體之間出現聯會與交換,這可能是引發物種不斷進行遺傳變異的根本機制。染色體的這種易變行為,可能是有性生殖的一種偶然而特殊的產物,但卻是物種變異的主要來源(當然由環境因子如射線引發的生殖細胞的突變也具有重要意義)。如果沒有有性生殖特有的變異機制,就不會有如今萬紫千紅的生命世界,也不會有我們今天的人類!
第三,任何一個物種的基因庫都有無限增加變異的趨勢,而它不可能容納無限的變異。也就是說,即便是一個數量相對穩定的群體,變異也將不斷地進行下去而導致基因庫的不斷擴增或變更,這主要源於減數分裂的內稟特性,也可能起因於物種在對環境(生物的和非生物的)緩慢適應過程中可獲得的一些可遺傳的變異。因此,任一物種的基因庫終將分裂(除非它提前消亡),新物種就此誕生,這個新誕生的物種又將進入新一輪的基因庫的變異累積與分裂的循環……。
因此,有性生殖物種的不斷分化不僅源於其特殊的基因保存方式,也根植於其基因庫的一種內稟特性之中,即物種不斷地重複著遺傳變異累積—基因庫分裂—遺傳變異累積的循環。這是有性生殖物種不斷分化的最重要的遺傳機制(謝平2013)。
發生在島嶼上的快速的隔離成種現象也說明物種基因庫的不斷變異性。達爾文曾注意到,生存在島嶼上的所有物種的數目比大陸上同樣面積上生存的生物要少,但其中大部分卻是本地的特有種。島嶼上大量新物種的出現不僅僅說明了隔離對成種的重要性,它同時也說明了任一物種的基因庫都在不停的變化,因為沒有後者(假如它的基因庫始終維持不變的話),即便某一物種的一些個體漂上了一座孤島,不論過多久它應該還是原來的種,與原來種群的個體還是能夠交配。當然,不同物種基因庫分裂的速率會有所不同,這種差異甚至還會很大。
具有諷刺意味的是,物種無限分化的趨勢被冠以了一些莫名其奇妙的機制。美國理論生物學家考夫曼說,“自主主體在不斷地走向新奇,如新的分子、新的形態、新的行為、新的組織,等等……適應於這樣一個自我構建系統的熱力學第四定律就是:它們要最大化它們的維數,即最大化下一時刻所發生事件的類型數”。這真是無稽之談!
生理學機制—氧化環境對物種分化的巨大貢獻
再來看看這樣一個事實:在現今的生命界,能夠在完全厭氧狀況下生存的生物僅限於最原始的生物類群—細菌和古菌,它們都是單細胞的原核生物,但大部分原核生物也都是好氧的。而真核生物都適應了好氧的生存方式,這與有氧代謝的能量利用效率遠高於厭氧發酵不無關係,因為後者僅為前者的1/4!
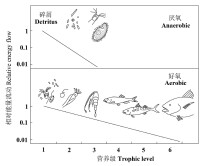
氧氣應該允許地球上更長食物鏈的存在
由於能量(利用)沿食物鏈逐級衰退,因此,食物鏈的能量利用效率越高(好氧),就允許越長的食物鏈存在。這樣,有氧食物鏈的出現就為生態系統食物網的複雜化(因此物種多樣分化)提供了重要的能量學基礎。一般認為,地球大氣圈的氧化歸功于海洋中的一類重要的原始光合生物—藍細菌,這些微小的單細胞生物,在數億年的生命演化歷程中,通過光合放氧以一種極為緩慢的方式將原始地球的厭氧大氣圈改造成了現在的氧化大氣圈。高效的有氧代謝體系的建立為真核生物的快速分化與繁榮奠定了最重要的生理學基礎。
生態學機制—生態位的不斷創造與細分
在地球上的生命誕生之初,哪裡會有那麼多的生態位來容納不斷增加的新物種呢?在過去數十億年間地球環境的演化歷史告訴我們,生命的進化不斷地創造或改造著自然界,譬如大氣圈的氧化為動植物的登陸創造了必需的生存條件,據說,臭氧層的出現可防止紫外傷害,這就為陸地生命的繁榮開闢了廣闊的空間,這其實是一次自然歷史上最偉大的生態位創造!
物種離不開食物與空間,即它們的生存資源,但地球表面的資源是有限的,而另一方面物種又具有不斷分化的內稟趨勢。這種資源的有限性與物種分化的無限性之間的對立就推動著地球表面的生態位的不斷細分。即便如此,地球也不可能無限地容納新物種,物種分化必定會被反饋平衡於一個有限的範圍內,因為當物種不斷分化時,總會有一些物種由於種群數量太小而使同種的個體間難以進行交配而導致滅絕,這樣分化與滅絕就達到了一個相對平衡。
從生態學的視角來看,自然界不斷重複著“物種分化—生態位創造—新物種再分化—生態位再創造......”的循環過程。即隨著物種的分化,可促進生存環境的改造與變化,這又導致生態位的增加,可為物種的生殖隔離創造新的條件,促進物種的再分化(細分)....如此循環往複。簡單地說,生命活動自身不斷地創造新的生態位,而新的生態位又為物種分化提供條件(創造了新的自然選擇目標)。另一方面,新舊物種間的競爭與適應也還會導致生態位的不斷細分。試問,如果不是這種生態位的不斷細分,何以能見到熱帶雨林的一棵大樹承載數以萬計物種的奇觀呢?
總的來說,物種分化與自然(生態位)創新是一種相互促進的正反饋關係,兩者呈現出一種無限分化或多樣化的趨勢,這似乎也與種群的無限增殖趨勢驚人地相似!從某種意義上來說,生命及其所形成的群落不僅是其自身進化歷史的產物,而且是物種生態關係演化的產物,還是氣候與地理環境的產物。
