bcl-2
bcl-2
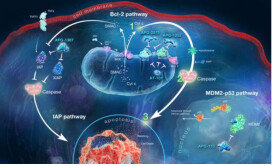
自從1972年Kerr提出細胞凋亡(apoptosis)的概念至今,人們對細胞凋亡現象進行了廣泛、深入的研究。但是,凋亡的分子和生化機制迄今尚未徹底明了,而已形成的初步認識大多源於對Bcl-2基因家族的研究。已知,凋亡進程可分為三個時相:誘導期,效應期和降解期。在誘導期,細胞接受各種信號從而引發各種不同的效應;進入效應期后,經過一些決定細胞命運(存活/死亡)的分子調控點,細胞進入不可逆的程序化死亡,這些調控分子包括一系列原癌基因和抑制癌基因的產生,其中Bcl-2家族起著決定性的作用;降解期則產生可見的凋亡現象。
Bcl-2基因(即B細胞淋巴瘤/白血病-2基因)是一種癌基因,它具有抑制凋亡的作用,並且近年來的一些研究已開始揭示這一作用的機制。目前已經發現的Bcl-2蛋白家族按功能可分為兩類,一類是像Bcl-2一樣具有抑制凋亡作用,如哺乳動物的Bcl-XL、Bcl-W、Mcl-1、A1、線蟲Ced-9、牛痘病毒E1B119kD等,而另一類具有促進凋亡作用,如Bax、Bcl-Xs、Bad、Bak、Bik/Nbk、Bid和Harakiri。
最初在血液淋巴細胞中發現Bcl-2能抑制細胞死亡,隨後陸續在其它一些細胞中也發現Bcl-2的這種作用。但近年來研究發現,除這些之外尚存在Bcl-2不敏感的凋亡途徑。
研究發現很多與Bcl-2同源性較高的基因,他們共同組成了龐大的Bcl-2家族。其中有的可以抑制凋亡,有的可以促進細胞凋亡。
又稱為抗凋亡蛋白(antiapoptotic proteins)有哺乳動物的Bcl-2,Bcl-XL、Bcl-W、Mcl-1、A1;線蟲Ced-9;牛痘病毒E1B119kD等
促凋亡蛋白,又稱為凋亡前體蛋白(proapoptotic proteins)Bax、Bcl-Xs、Bad、Bak、Bik/Nbk、Bid和Harakiri。這些又可以分為兩類
● ● 僅含BH(Bcl-2 homology)3結構同源域的:Bik,Bid,Bad,Bim,Bmf等。
● ● 含有多個BH同源區域的成員:Bax, Bak,Bcl-xs,Mtd等。
Bcl-2可以抑制由多種細胞毒因素所引起的細胞死亡。Bcl-2的過度表達能增強所觀察細胞對大多數細胞毒素的抵抗性。這一發現使人們認識到凋亡的各種信號轉導途徑有一個共同的通路或交匯點,而該通路或交匯點受Bcl-2調節。實驗表明,Bcl-2可增強細胞對大多數DNA損傷因子的抵抗性,抑制大多數化療藥物所引起的靶細胞凋亡,但其本身並不能抑制這些因素對細胞的損傷;同樣地,它也不能促進DNA修復。p53蛋白是DNA損傷的一個分子感測器,已證實Bcl-2能抑制p53介導的凋亡,但不能抑制p53向核內轉位或者p53介導的生長停滯,可能Bcl-2的作用是在DNA損傷后,阻止激活凋亡機制的信號到達其靶分子;在細胞毒性T細胞中,由顆粒酶B激活Caspases家族的一個或多個半胱氨酸蛋白酶所誘導的凋亡不依賴於Bcl-2,顆粒酶B可能僅作用於凋亡路徑中Bcl-2調節位點的下游。以上結果表明在凋亡途徑中Bcl-2的作用位點是在信號分子和效應蛋白酶之間的位置。
Bcl-2抑制細胞凋亡可能與其以下幾種作用有關:
① 細胞催抗氧化作用以往研究表明,在去除生長因子所誘發的凋亡中,活性氧損傷(ROS)是促發細胞死亡的主要誘因:Bcl-2的過度表達可減少氧自由基的產生和脂質過氧化物的形成。提示Cai等的實驗表明Bcl-2的抗氧化作用是間接的,即可能在於抑制超氧陰離子的產生而不是直接清除活性氧。細胞色素c(cytochrome c, Cyt c)作為呼吸鏈中重要的電子傳遞體,它從線粒體內膜上的釋放會阻斷電子向下游傳遞體危及呼吸鏈的功能並導致超氧陰離子的加速產生;而Bcl-2可以抑制Cyt c的釋放,從而抑制了超氧陰離子的產生。此外,Bcl-2還可以增加胞內的谷胱甘肽(GSH)等抗氧化劑,升高NAD/NADH的比值,抑制和凋亡相聯繫的GSH降低,促進GSH進入核內,從而影響細胞的氧化還原狀態。但也有證據表明,在沒有自由基的情況下Bcl-2也能抑制凋亡。因此,Bcl-2的性質遠非僅僅是一種抗氧化劑。
② 抑制鈣離子跨膜流動 Lam等曾報道鈣泵特異性抑製劑Thapsigargen(TG)誘導的WEH17.2等細胞的凋亡可被Bcl-2所抑制,其原因是Bcl-2抑制了Ca2+的跨膜流動,提 示Bcl-2可通過調節胞內鈣離子濃度來調節凋亡。然而Wei等在神經元細胞GT1-7中發現Bcl-2中發現Bcl-2可抑制由TG所誘導的凋亡,但並不能抑制TG誘導的胞內鈣離子濃度升高;同時Jason等在中國倉鼠卵巢細胞5AHSmyc中去除細胞內貯存Ca2+后發現過度表達的Bcl-2仍可抑制凋亡,提示Bcl-2對胞內Ca2+濃度的調節作用也只能是其抑制凋亡的機制之一。
③-1 離子通道蛋白 體外實驗證實,Bcl-2、Bcl-X1和Bax能在線粒體上形成離子通道,提示它們可能參與調節一些與凋亡有關的細胞現象,如線粒體通透性改變(巨孔形成)和線粒體釋放凋亡蛋白本科激活因子——Cyt c和凋亡誘導因子(AIF)。過度表達的Bcl-2能抑制線粒體通透性改變,並影響巨孔的形成,從而抑制凋亡。但也有實驗發現即使線粒體通透性未改變,Cyt c等凋亡激蛋白也可以釋放到胞液中從而誘發細胞凋亡,因此這種觀點還有待進一步證實。
在所觀察的細胞系及多種情況下,凋亡的發生都伴有Cyt c從線粒體釋放,繼而伴有Caspases激活,而活化的Caspases降解底物后,其蛋白降解產物又引起線粒體通透性改變,並最終導致凋亡發生。Yang等與Kluck等發現Bcl-2可以通過抑制線粒體釋放Cyt c來抑制凋亡,繼而Rosse等和Zhivotovsky等用兩種不同的方法證明了即使Cyt c已經釋放入胞漿,Bcl-2仍能延遲細胞死亡,這是由於Bcl-2抑制了活化的Caspases對線粒體釋放的上游和下游都有Bcl-2調節凋亡的作用點。
AIF作為一種線粒體來源的效應蛋白酶,在分離在胞核中能誘導凋亡,同時還能激發線粒體通透性改變,誘導線粒體釋放Cyt c和Caspase-9,並在體外切割和激活其底物——半胱氨酸蛋白酶CPP32。體外研究發現,Bcl-2過度表達可能阻止AIF從分離的線粒體上釋放,但它並能影響AIF的產生,同時也不能影響AIF促凋亡的功能。
Bcl-2可與Bcl-2家族的Bcl-X1、Bcl-Xs、Bax、Bcl-2、Bad和Mc1-1形成同源的蛋白二聚體,而特定的蛋白二聚體則可作為在細胞死亡信號通路上的分子開關。例如,Bcl-2可與促凋亡Bax形成二聚體,如果Bax相對量高於Bcl-2,則Bax同二聚體的數量增多,從而促進細胞死亡;而如果Bcl-2相對量高於Bax,則促進形成Bcl-2/Bax異二聚體,並使Bcl-2同二聚體的量增多,從而抑制細胞死亡;而促凋亡分子和Bad和Bcl-Ss則競爭結合細胞死亡抑制分子如Bcl-X1或Bcl-2,由此替換了Bax並促進Bax同二聚體形成,誘導凋亡。
另外,Bcl-2還可與Bcl-2家族外的11種蛋白質結合,包括蛋白激酶Raf-1、蛋白磷酸酶Calcineurin、GTP酶、R-Ras和H-Ras、p53-BP2、朊蛋白Pr-1、Ced-4、BAG-1、Nip-1、Nip-2和Nip-3,並由此實現其抗凋亡作用。
Bcl-2抑制凋亡的作用並不是萬能的,例如在細胞毒T細胞殺傷作用中,特定的淋巴因子撤離以及某些TNF受體家族介導的凋亡途徑中Bcl-2不能抑制凋亡,其原因右能是這些凋亡誘導劑作用於Bcl-2下游或是獨立於Bcl-2的其它凋亡路徑。最近Scaffidi等發現,Jurkat等細胞(Ⅱ型細胞)中Bcl-2可抑制TNF受體家族中的CD95(Apo-1/Fas)信號通路介導的凋亡,而在SKW6.4等細胞(Ⅰ型細胞)中Bcl-2則不能抑制這種信號通路介導的凋亡。其結果表明,凋亡途徑對Bcl-2敏感與否可能與細胞類型有關。進一步研究發現,這種差異取決於線粒體是否參與這種凋亡途徑,在Ⅰ型 細胞中線粒體則未參與該凋亡途徑。因此,Bcl-2可能只抑制依賴於線粒體的凋亡途徑。
更進一步研究發現,來源於牛痘病毒的細胞因子應答修飾體A(crmA)是一種絲氨酸蛋白酶抑製劑(serpin)類分子,能抑制一些Caspases的功能。而來源於桿狀病毒的P35蛋白也能抑制一些Caspases的功能,而且它比CrmA具有更廣泛的Caspases抑制特性。在組織培養體系中或轉基因鼠的T淋巴細胞中CrmA表達能抑制CD95(Fas/Apo-1)和p55TNF受體Ⅰ誘發的凋亡,但對另一些諸如生長因子撤除,DNA損傷等細胞毒因素引起的凋亡則無能為力,而這些由細胞毒因素引起的凋亡現象則可被Bcl-2及其同系物所抑制,但是Bcl-2又不能抑制由CD95(Fas/Apo-1)和TNF受體介導的凋亡。由細胞毒因素及TNF受體家族介導的凋亡都可有效地被p35所抑制,提示這些通路都依賴於Caspases的激活。上述結果表明,在細胞中不同的凋亡激活途徑同時存在,其一是需要對CrmA敏感的Caspases參與但又不能被Bcl-2所抑制;而另一個則是Bcl-2可抑制的凋亡途徑,其中Caspases對p35敏感,但對CrmA則不敏感。
目前Bcl-2是凋亡分子機制研究的主要靶分子。隨著對Bcl-2以及凋亡本身研究的日漸深入,Bcl-2的作用機制和凋亡的分子機制最終被闡明,從而提高對與凋亡有關的疾病的認識和診治水平。
B-cell CLL/lymphoma 2
基本信息
- 中文名
- B淋巴細胞瘤-2基因
- 外文名
- B-cell lymphoma-2
- 屬性
- 癌基因之一
- 領域
- 細胞凋亡研究