有孔蟲目
有孔蟲目

有孔蟲目

有孔蟲目
有孔蟲的生殖方式為有性生殖的配子母體世代與無性生殖的裂殖體世代交替進行。主要生活於正常海洋環境中,淡水種極少。多數種底棲生活,浮遊性種類較少。有孔蟲是微雜食性動物,也有腐食性的。
各個地質時期的有孔蟲化石常被用作確定地質年代的標準化石和古沉積環境的指相化石,特別是被用於恢復新生代的古氣候、古海洋環境(如古水深、古溫度、古鹽度、古海流、古水團)以及估計沉積率等等,有助於石油勘探。
有孔蟲目動物的整個細胞質團由含各種不同礦物成分的殼體包裹著。原生質分外質、內質兩層。外質色淺透明,可通過口孔或殼壁孔伸延殼外,成細線狀或粒網狀偽足,具有運動、攝食、消化、呼吸、排泄、分泌、造殼等功能。內質色深,有各種細胞器,如細胞核、環片、線粒體、微體、高爾基器、溶酶體、微管、黃色素體、共生體等。
有孔蟲胞核的數目因種類而異,有單核的或多核的。多核種類又分異型核的和同型核的兩類。在生殖發育階段,也存在著二型核現象:有性生殖世代(顯球型)是單核的;無性生殖世代(微球型)則為多核的。
殼體依殼壁組成成分及微細構造,可分為以下幾種基本類型:①假幾丁質殼,最原始類型,薄而易變形(如網足蟲亞目);②膠結質殼,殼壁由自身分泌的物質膠結外來物質而成(如編織蟲亞目);③鈣質殼,主要由自身分泌的碳酸鈣組成,依殼壁的結晶體和微細構造,可分為微粒狀無孔殼、似瓷質無孔殼(粟蟲亞目)、鈣質透明有孔單層放射狀殼(瓶蟲亞目)、鈣質透明有孔多層粒狀殼(輪形蟲亞目)、鈣質透明放射微粒狀殼(包旋蟲亞目)、鈣質透明單晶殼(旋蟲亞目)、鈣質骨針殼(卡特蟲亞目);④硅質殼,殼壁成分為二氧化硅(如硅室蟲亞目)。
殼體有單房室、雙房室或多房室。房室間的隔板有孔相通,有孔蟲即以此特徵而得名。單房室類群多數呈近球形或平旋管狀;多房室類群的房室排列多數為螺旋式,其次為平旋、雙列、盤狀、粟蟲式旋卷、單列等,還有混合排列,如從平旋到單列或雙列,從雙列到單列,從螺旋到三列等形式。
繁殖方式
有孔蟲目動物的生殖方式為有性生殖的配子母體世代與無性生殖的裂殖體世代交替進行。有孔蟲目生物的生命周期包括雙倍體及單倍體產生的交替,雖然兩者形態上大部分相近。單倍體或產配子體(gamont)初期有一個細胞核,及後分裂成為很多生殖細胞,通常有兩個鞭毛。雙倍體或裂殖體(schizont)為多核細胞(multinucleate),經過減數分裂後分裂製成新的產配子體。有性別的世代在水底進行多重無性生殖並不罕見。
成熟的裂殖體減數分裂,行無性裂配生殖,形成許多配子母體。配子母體成熟后,產生許多帶有2根鞭毛的配子,每兩個配子融合成合子,行有性配子生殖。從合子發育成裂殖體為二倍體期。從減數分裂進行裂殖生殖至配子母體產生配子,則為單倍體期。
有性生殖配子母體世代的初房較大,稱顯球型殼;無性生殖體的殼體初房則較小,稱微球型殼,這就是有孔蟲殼體的二型現象。這種二型現象僅見於底棲種類,浮遊有孔蟲無此現象。有些底棲有孔蟲配子的形態與鞭毛數目是有變化的,例如旋蟲、帽貝蟲的配子為變形蟲狀,不帶鞭毛,有的種具3根鞭毛,還有如玫瑰蟲在配子母體世代發育1個浮室以產生配子。有孔蟲經世代交替完成整個生活史所需時間一般為1~2年或更長時間,但也有隻需半個月左右的。
活動及進食
假足的作用在於移動、固定及捕捉食物,包括細少生物例如硅藻(diatom)或細菌。部分形態為以內共生體(endosymbiont)的單細胞藻類,有多種多樣的世系如綠藻(greenalga)、紅藻(redalga)、金藻(goldenalga)、硅藻及溝鞭藻類(dinoflagellate)。部分有孔蟲門生物為盜食質體(kleptoplasty),保留攝取藻類的葉綠體去進行光合作用。
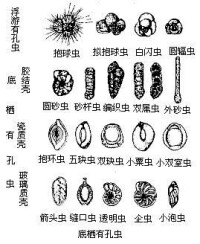
有孔蟲目
底棲有孔蟲自潮間帶至深海盆均有分佈,分佈最深記錄是在千島堪察加海溝(10687米)。在東海,底棲有孔蟲種類和數量的分佈趨勢是:河口區稀少,自內陸架向中、外陸架迅速增多;越過陸坡向沖繩海槽區,種類逐漸減少,數量則顯著減少。浮遊有孔蟲一般是近岸稀少,越遠離海岸,種類和數量越增加。浮遊有孔蟲有晝夜垂直移動現象。
鹽度對有孔蟲的影響很顯著,在正常海洋鹽度區,底棲有孔蟲種類多、數量大;在低鹽的河口區、瀉湖和沼澤區,有孔蟲種類貧乏,在高碳酸鈣、超高鹽的潟湖中,較常見的是粟蟲科的種類(如三浹蟲)。
溫度是影響有孔蟲分佈的重要因素。在熱帶,有孔蟲的生殖臨界溫度遠比溫、寒帶的狹窄。熱帶-亞熱帶有孔蟲的種類與數量遠比溫、寒帶繁多。浮遊有孔蟲的緯度分佈和底棲有孔蟲的深度分佈,明顯受到水溫的制約,生活於熱帶深海盆的底棲冷水種,在高緯度則可分佈於較淺海底。溫度還可以影響有孔蟲殼體的形態構造,如厚壁新方球蟲在兩極區,殼體左旋;在溫暖海域,殼體則為右旋。
大洋環流控制著浮遊有孔蟲的地理分佈。布拉德紹於1959年所劃分北太平洋的4個浮遊有孔蟲區系(亞北極區系、過渡帶區系、中央區系、西赤道中央區系)與北太平洋幾個大水團的分佈範圍適相對應。馬維驊1977年把世界大洋浮遊有孔蟲劃分為3個生物地理區(環熱帶區、北冷水區、南冷水區)、5個省(北極與南極省、亞北極與亞南極省、南北過渡省、亞熱帶省、熱帶省)。大洋環流的界線與浮遊有孔蟲區系界線十分一致。
底質對有孔蟲有明顯影響,在富含有機質的粉砂和泥質底質中,有孔蟲的數量常較多,且多數是薄殼或細長類型。在貧營養的砂或砂礫底質中,有孔蟲數量常較少,且多數為厚殼透鏡形或紡錘形類型。生活在硬底質如岩石、多貝殼或多海藻環境中的有孔蟲,多數是便於貼附或附著的殼形。碳酸鈣在暖水區的溶解度比在冷水區小,所以厚鈣質殼有孔蟲、有孔蟲石灰岩和有孔蟲軟泥都出現在低緯度區。碳酸鈣溶解度隨著水深增加而增大。在碳酸鈣溶解度變化最大的溶躍面以下,鈣質有孔蟲殼大多數被溶解,因而深海有孔蟲動物群是以膠結種類佔優勢。

二疊紀為古生代有孔蟲繁盛時期,以內卷蟲等占絕對優勢。似瓷鈣質無孔有孔蟲,也在這一時期開始出現。晚古生代發生了劇烈的構造運動和大陸運動,海退大幅度改變了水圈條件,導致三疊紀有孔蟲種類發展緩慢。
到了侏羅紀,氣候轉暖而平穩、海浸廣泛,世界有和暖的邊緣海和環赤道暖流系。這一時期堪稱有孔蟲盛期,各種類型的鈣質和膠結底棲有孔蟲十分豐盛,並出現文石質殼的羅伯特蟲類群,浮遊有孔蟲在此期開始出現。到白堊紀,浮遊有孔蟲更為豐盛,成為白堊紀優異的標準化石。
但到了白堊紀晚期,大多數白堊紀浮遊有孔蟲和淺水底棲有孔蟲絕滅,而大多數深水底棲有孔蟲仍延續出現。這一事實提示了當時海洋環境的劇烈變化僅局限於200~300米的上面水層。
第三紀初期古新世一些白堊紀浮遊有孔蟲類群如輪孔蟲科、截球蟲科、沙科蟲科已全部絕滅,而抱球蟲科成為最主要的浮遊類群。同時,還出現漢京蟲科、圓輻蟲科等類群。始新世大型鈣質底棲有孔蟲,如貨幣蟲科、鱗環蟲科、蜂巢蟲科等都很豐盛,形成石灰岩。中新世有孔蟲基本具備了現代有孔蟲的屬種面貌。
20世紀60年代以來,世界各國對各大洋有孔蟲的調查研究,“深海鑽探計劃”(DSDP)的成功,電子顯微鏡和電子探針等新技術的應用,細胞學、生理學、生物化學、生態學等現代生物學概念以及新的研究手段、新的資料分析方法的引入,促使對有孔蟲的研究向新的高度發展。
地層測量
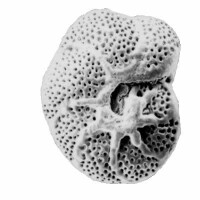
有孔蟲目
古代環境的重建
石灰質的有孔蟲目生物化石由其生活的古代海洋元素組成。所以其化石對古氣候學(paleoclimatology)及古海洋學(paleoceanography)十分有用。她們一方面可以利用測定氧氣中穩定同位素(stableisotope)的比例去重組古代氣候,另一方面也可以利用測定碳中穩定同位素(stableisotope)的比例去重組碳循環的歷史及古海洋生產力。浮遊有孔蟲目生物化石記錄的地理模式亦可以用作重組古代洋流情況。因為部分有孔蟲目生物只可以在特定環境中找到,它們可以用作找出每種古代海洋沉積物所沉積的環境。
氣候及環境的生物指示劑
基於對於生物地層學有價值的同樣原因,活有孔蟲目生物的集合物亦被利用作為海岸地區的生物指示劑(bioindicators),包括指示出珊瑚礁的健康程度。因為碳酸鈣易於溶在酸性環境的影響,有孔蟲目生物可能特別受到氣候轉變及海洋酸化(oceanacidification)的影響。
原生動物門肉鞭動物亞門根足亞綱抱球蟲科海洋環境珊瑚礁生理學生物化學生態學深海鑽探白堊紀侏羅紀同位素二疊紀寒武紀奧陶紀
1.http://bio.biox.cn/Biology/200701/20070106220733_24482.shtml
1.http://www.kpzs.net/animal/animal/787/200710/308057.html
2.http://www.wanfangdata.com.cn/qikan/periodical.Articles/DCXZZ/dcxz2007/0704/070402.htm