刺胞動物門
刺胞動物門
刺胞動物徠門(Cnidaria)又稱刺細胞動物門,過去稱為腔腸動物門(Coelenterata),因為它的含義適用於刺胞動物及櫛水母動物,所以現多已廢棄不用。
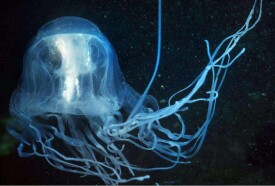
刺胞動物門(Cnidaria)又稱刺細胞動物門,過去稱為腔腸動物門(Coelenterata),因為它的含義適用於刺胞動物及櫛水母動物,所以現多已廢棄不用。刺胞動物體呈輻射或兩輻射對稱,僅具二胚層,是最原始的後生動物。體壁由外胚層、內胚層和中膠層組成。內胚層圍成身體的整個內腔稱消化循環腔,腔腸一端為口,他端閉塞,無肛門。體壁中有刺細胞。腔腸動物的骨骼主要為外骨骼,具有支持和保護功能。多由幾丁質、角質和石灰質構成。在很多珊瑚蟲中具有骨針或骨軸,它們存在於中膠層,或突出於體表面。有無性和有性兩種生殖方式,並往往出現在同一種的生活史中,即水螅型世代用無性出芽生殖方式產生水母型世代,而水母型個體脫離母體,長大成熟以後,又以有性生殖方式產生水螅型個體,兩個世代互相交替完成整個生活史。約1萬種,可分為:水螅蟲綱、缽水母綱和珊瑚蟲綱3綱,一般認為水螅蟲綱是最原始的。絕大部分海產,只有少數種類產於淡水,以熱帶和亞熱帶海洋的淺水區最豐富。較小種類可作魚類食餌,珊瑚骨骼(如紅珊瑚)可作工藝品,古珊瑚和現代珊瑚可形成儲油層,海邊的珊瑚礁可作天然海堤;海蜇鹽漬后成為食品,有些種類可作藥用;有些浮遊水母類可作為海流指示生物;但是,有些大型水母(如霞水母、根口水母)在大量出現時,會阻塞或破壞漁網,有的海葵混於漁獲物中,人食后中毒致死。刺胞動物除極少數種類為淡水生活外,絕大多數種均為海洋生活,多數在淺海,少數為深海種,現存約11000種,分為3個綱:水螅綱(Hydrozoa)、缽水母綱(Scyphozoa)、珊瑚綱(Anthozoa)。
刺胞動物很少能做主動的移位運動,運動的能力是很有限的,特別是水螅型。運動是由表皮肌肉細胞中肌原纖維的收縮所引起。例如水螅的身體可做伸縮運動,伸展時體長可達15~20mm,收縮時體長僅0.5mm,這種伸縮是爆髮式的,每5~10分鐘左右爆發一次,主要是由外皮肌細胞的縱行肌原纖維的收縮所引起。身體一側的肌原纖維的收縮可引起身體的彎曲。有時靠彎曲身體及觸手並與基盤的交替附著而做翻斛斗式的運動。基盤處粘細胞可分泌大量的氣泡,可使水螅在水面上做短暫的漂浮。缽水母類和珊瑚類的肌原纖維已與表皮細胞分離形成獨立的一層肌纖維,例如水母的肌原纖維在下傘面及傘緣形成薄薄的一層肌肉環,有的被輻管分離成片狀,其纖維有橫紋,它們做有規律的收縮,使傘面有節奏的收縮運動。當傘緣收縮時,傘緣內的水被噴出,由於反作用力而推動水母體向上運動,當傘及傘緣肌肉舒張,被壓縮的中膠層的彈性,使傘又恢復了原形,水又重新進入傘緣內,身體下沉,但由於收縮比舒張要快,所以水母還是可以向上垂直運動。水母類的垂直運動在有緣膜的水母可以看的更為清楚。一旦肌肉停止收縮,水母就會自然下沉。其水平方向的運動多是被動的,多是由於水流及風力所推動。又例如在珊瑚類,海葵的肌原纖維在隔膜上形成發達的牽縮肌,它的收縮非常有力,所以當海葵固著在岩石上是很難將它取下的。
刺胞動物都是肉食性的,以浮游生物,小的甲殼類、多毛類甚至小的魚類為食。由於食物的機械刺激和化學刺激,引起水螅類動物伸長觸手,並放出刺絲囊以纏繞、麻痹、毒殺捕獲物,再將食物送入口中。口區腺細胞分泌的粘液有利於食物的吞咽,食物進入胃腔后,胃層的腺細胞開始分泌蛋白酶,分解、消化食物使之形成許多多肽,同時在胃腔中由於營養肌肉細胞的鞭毛運動,食物得以混合與推動。經這種細胞外消化之後,開始細胞內的消化過程,營養肌肉細胞的偽足吞噬食物顆粒,在細胞內形成大量的食物泡,經過酸性及鹼性的化學過程之後,營養物質由細胞的擴散作用輸送到全身。缽水母類及珊瑚類胃腔結構比較複雜。缽水母的胃腔中有各種輻管及環管,胃囊中有內胚層起源的胃絲;珊瑚類的胃腔被許多隔膜分隔成許多小室,隔膜上有隔膜絲。胃絲及隔膜絲中含有大量的刺細胞及腺細胞,它們是將食物吞入胃腔之後才殺死及消化。消化后的營養物通過各種管道輸送到全身,未消化的食物殘渣仍由口排出。糖元及脂肪是腔腸動物的主要貯存物。刺胞動物中許多種類,特別是海洋中的造礁珊瑚類,體內均有共生的藻類,如動物黃藻、腰鞭毛藻之類。藻類能進行光合作用,產生甘油、脂肪、糖及脯氨酸等,並提供給刺胞動物作為其補充營養。這將在珊瑚綱中做進一步的敘述。
刺胞動物沒有專門的呼吸及排泄器官,由於身體是由兩層細胞圍繞胃循環腔所組成,並通過口使胃腔與外界相通,實際上體壁的兩層細胞均與外界環境接觸,所以呼吸與排泄作用可以由體壁細胞直接獨立進行。出、入口及胃腔的水流可以攜帶入新鮮的氧氣,並帶走代謝產物。刺胞動物的主要含氮廢物是氨。
刺胞動物是最早出現神經結構的多細胞動物。神經結構原始,是由雙極神經元、多極神經元及神經纖維聯合成神經叢或神經網,一般位於外表皮細胞基部肌肉層之外,呈網狀分佈,故稱網狀神經系統(netnervous system)。由於神經細胞可向各個方向傳導,也稱為漫散神經系統(diffusenervous system)。水螅就只有一個神經網,但大多數腔腸動物除了表皮的神經網之外,在胃層的基部也有一個神經網,這兩個神經網或是完全獨立的,或有纖維相聯。1971年Westfall等首先證明了水螅及其他腔腸動物在神經元之間、神經元與效應器(表皮肌肉細胞、刺細胞、腺細胞等)之間也存在著突觸傳遞(synaptic transmission)。即神經衝動的傳導是由一個神經元的軸突,傳遞給另一個神經元的樹突。軸突的末端具有突觸小泡(synaptic vesicle),樹突的末端沒有。傳導時,突觸小泡釋放乙醯膽鹼引起后一個神經元的興奮,衝動傳出以後,乙醯膽鹼被神經末稍表面的膽鹼酯酶水解成膽鹼及乙酸而解除了激活作用。以後又發現腔腸動物的突觸傳導分為兩種類型。一類突觸是對稱性的傳導(symmetrical),即兩個或多個神經末端有突觸小泡,因此衝動可以同時向兩個或幾個方向傳遞,這種傳導也叫非極性的傳導(nonpolarized transmission);另一類突觸是不對稱的(asymmetrical),即神經細胞只有一個末端具突觸小泡,神經衝動只能向一個方向傳導,因此也稱為有極性的傳導(polarized transmission)。最近的研究證明刺胞動物的非極性傳導是很普遍的。也有人認為刺胞動物的兩個神經網一個是由多極神經元形成的非極性傳導,也就是慢傳導系統;另一個是由雙極神經元形成的極性傳導,也就是快傳導系統。刺胞動物的神經元可以有2個、3個或多個長的神經突起,這些突起或與感官相聯、或與效應器相聯、或與其他神經元相聯、或者是同一個神經元同時與感官與效應器相聯,這種多樣性說明了神經細胞傳導的原始性。在生理意義上代表了高等動物所具有的感覺神經、運動神經及中樞神經的起始階段。此外,刺胞動物的非神經傳導,即上皮傳導也是很常見的,例如刺細胞、表皮肌肉細胞可以完全獨立於神經元而被自身所控制。水母型的神經結構比水螅型複雜,例如水螅水母,除了傘部的神經網之外,傘緣的上皮神經細胞分別在傘緣的上、下面集中形成兩個神經環。下面(或稱外面)的一個神經環更發達。缽水母類的水母型多數沒有這種神經環,而是在傘緣集中形成4~8個神經節。水母型個體均能表現出一種有節奏的收縮運動,這是由於水母的神經結構每隔一定時間能自發地產生動作電位,並經過神經傳導引起整個身體的收縮。研究已經證明這種自發的動作電位是由起博點的神經元所引起。起博點神經元在水螅水母存在於外神經環中,在缽水母類存在於感官神經節中。而且每一個神經節(一般4~8個)都有起博神經元,它是水母類有節奏博動的中心。一個水母體似乎有一個起博點就足以引起身體有節奏的收縮運動,但實驗證明多個起博點比僅有一個起博點能使收縮的節奏更有規律、更有保障、使間隔的時間也較短。腔腸動物的水螅型體沒有明顯的感官,其感覺細胞可分佈全身,但觸手、口區較為豐富。水母型個體,在傘緣具有豐富的感覺細胞或感覺器官。感覺細胞的細胞體都是具纖毛的。感覺器官包括眼點及平衡囊。眼點是由感覺細胞構成的杯狀物,內有色素顆粒分佈,它對光線或有正趨性,即有光時游向水面。或對光有負趨性,即有光時下沉水底,無光或弱光時浮向水面。平衡囊在水螅水母結構簡單,在緣膜基部或下傘面神經環(即外神經環)處形成一個小囊,囊的內壁有感覺細胞,細胞上也有纖毛。囊的底部有一鈣質結石。它是一種重力感受器。當傘緣傾斜時,結石與感覺細胞的纖毛接觸,並刺激纖毛細胞以產生動作電位,抑制了該側肌肉纖維的收縮,通過肌肉收縮再調整身體使恢復平衡位置。缽水母類的平衡囊結構更複雜,將在有關節中敘述
無性生殖及有性生殖在腔腸動物中都是很普遍的。無性生殖的主要形式是出芽生殖,特別是在水螅型更為常見。例如水螅出芽時從身體的靠基部通過體壁及胃腔的向外突出,再長出觸手與口即形成芽體,以後芽體與母體分離即形成新的個體。但在藪枝螅(Obelia)所形成的芽體與母體不分離則形成了群體。其次,也可以通過分裂方式進行無性生殖,主要發生在水螅型,例如海葵可以縱分裂,缽水母的幼體以橫分裂進行無性生殖。此外水螅型一般具有很強的再生能力,例如,將水螅切成數段,條件合宜時每段都可以再生成一個新的個體,再生時口與反口端的極性不變,但口端的再生速度比反口端迅速。所以水螅型的再生現象也被認為是無性生殖的一種方式。有些生殖出現在多數水螅型及所有的水母型,除少數種類例如水螅為雌雄同體之外,絕大多數種類為雌雄異體(或異群體)。生殖細胞來源於間細胞,然後遷移到固定的位置上形成生殖腺。水螅蟲綱的動物生殖腺來源於表皮層,例如水螅,水螅水母的生殖腺位於放射管下或垂唇周圍,但都是來源於表皮層細胞。缽水母綱的生殖細胞起源於胃層位於胃囊低部。珊瑚綱的生殖細胞是由胃腔中的隔膜上發生,也是內胚層來源。腔腸動物只有生殖腺,沒有出現生殖導管及生殖附屬腺,生殖細胞成熟之後由口排出,或由體壁破裂而釋放。受精作用依不同的種而異,或在體外海水中進行,或在垂管的表面,或在胃腔內生殖腺的部位進行。卵裂是完全的,形成中空的囊胚,經移入法或內陷法形成原腸胚,結果形成兩層細胞,即兩個胚層,內部成團的細胞為內胚層,將來形成成體的胃層,外表的一層為外胚層,將來形成成體的表皮層。實心的原腸胚迅速延長,體表出現纖毛,形成了自由游泳的浮浪幼蟲(planula)。浮浪幼蟲早期沒有口及胃腔,游泳一段時間之後,固著在水草、岩石或其他物體上發育成水螅型體,或再經過出芽生殖形成群體。淡水生活的水螅沒有幼蟲期,其受精卵直接發育。
| 身體呈輻射對稱,有的為兩輻射對稱 |
| 兩胚層和原始消化腔 |
| 細胞出現原始的組織分化 |
| 網狀神經系統(擴散或散漫神經系統) |
| 特有的刺細胞 |
| 水螅型和水母型 |
| 有性和無性生殖,有世代交替現象,海產種類有浮浪幼蟲期 |
| 輻射對稱與兩輻射對稱 | |
| 多孔動物的體型多數是不對稱的。從腔腸動物開始,體型有了固定的對稱形式。本門動物一般為輻射對稱(radial symmetry)。即大多數刺胞動物,通過其體內的中央軸(從口面到反口面)有許多個切面可以把身體分為2個相等的部分。這是一種原始的低級的對稱形式。這種對稱只有上、下之分,沒有前後左右之分,只適應於在水中營固著的或漂浮的生活。利用其輻射對稱的器官從周圍環境中攝取食物或感受刺激。在刺胞動物中有些種類已由輻射對稱發展為兩輻射對稱(biradial symmetry),即通過身體的中央軸,只有兩個切面可以把身體分為相等的兩部分。這是介於輻射對稱和兩側對稱的一種中間形式。 |
| 兩胚層與原始消化腔 | |
| 多孔動物雖然具有二胚層,但從發生來看,它與其他後生動物不同,因此一般只稱為二層細胞。刺胞動物才是具有真正二胚層(內、外胚層)的動物。在二胚層之間有由內、外胚層細胞分泌的中膠層。由內外胚層細胞所圍成的體內的腔,即胚胎髮育中的原腸腔。它與海綿的中央腔不同,具有消化的功能,可以行細胞外及細胞內消化。因此,可以說從這類動物開始有了消化腔。這種消化腔又兼有循環的作用,它能將消化后的營養物質輸送到身體各部分,所以又稱為消化循環腔(gastrovascular cavity)。有口、沒有肛門,消化后的殘渣仍由口排出。它的口有攝食和排遣的功能。口即為胚胎髮育時的原口,與高等動物比較,可以說腔腸動物相當於處在原腸胚階段。刺胞動物的表皮層與胃層之間為中膠層(mesoglea),水螅型體內的中膠層不發達,一般為很薄的一層,其中很少有細胞分佈。水母型體內中膠層十分發達,佔據了身體的幾乎整個厚度,其中含有纖維及少量來源於外胚層的細胞。中膠層中的主要成分是水分,其中含有極少量的蛋白質及多糖,其濃度一股低於1%,其滲透濃度比海水還低,但由於其中含有少量的硫酸鹽和其他離子(硫酸鈣溶液比等滲的氯化鈣溶液更濃),所以它的總滲透濃度和海水相近。一般情況中膠層中含有機物為5%,也有人用細指海葵(Metridium)做實驗,發現其中膠層中含有8%的蛋白質和1%的多糖,並以膠原蛋白質的形式存在,這種物質具有很強的彈性和粘合力。脊椎動物肌腱中的膠原纖維只能延伸到自身長度的10~20%,刺胞動物中膠層中的膠原纖維卻能伸長自身長度的三倍(橡膠是四倍)。所以中膠層不僅使刺胞動物可以伸縮及保持體形,在一定意義上也是刺胞動物的一種骨骼類型。 |
| 細胞組織的分化 | |
| 海綿動物主要是有細胞分化。刺胞動物不僅有細胞分化,而且開始分化出簡單的組織。間細胞是小型未分化的胚胎性細胞,可分化成皮肌細胞、刺細胞、生殖細胞等。腺細胞(glandcell)是一種具分泌能力的上皮細胞。在水螅的基盤處及觸手的表皮層中腺細胞特別發達,它的分泌物可以幫助水螅體附著及捕食等。在具有骨骼的種類,例如水螅型群體及珊瑚類,其腺細胞可以分泌大量的角質或鈣質以形成幾丁質的圍鞘或鈣質外骨骼。胃層中也含有大量的腺細胞,細胞內含有大量的分泌顆粒,它可以轉化成消化酶,進行食物的細胞外消化。間細胞(interstitialcell)位於上皮細胞之間,靠近中膠層處,是一些小型、圓形的細胞,單獨或成堆分佈,具大的細胞核,它是體內一種未分化的細胞,由它可以轉化成生殖細胞、刺細胞、腺細胞等其他類型的細胞。感覺細胞(sensory cell)細胞體長形,垂直於體表,在口區及觸手處很豐富,細胞基部具有很多神經突起,端部具感覺毛,感受各種刺激,然後經神經突起作用於效應器或細胞。 |
| 水螅型與水母型 | |
| 刺胞動物的體型具有兩種基本形態,即營固著生活的水螅型及營漂浮生活的水母型。在水螅蟲綱,原始的種類水螅型與水母型交替出現在生活史中,無性生殖階段表現為水螅型,有性生殖階段表現為水母型,大多數水螅蟲綱其水螅型比較發達,特別是群體生活的種類。在缽水母類水母型發達,水螅型不發達或完全消失。珊瑚蟲類只有水螅型體型、水母型已不復存在了。水螅型呈單體或群體,例如水螅(Hydra),這是一種池塘中常見的單體生活的動物,身體呈筒形或管形,長約5mm左右,從口到基盤的體軸名為口—反口軸(oralaboralaxis)口端遊離、頂端有口,口的周圍有一圈觸手(tentacles),觸手的數目水螅為5~10個,其他種類因種而異。水螅的反口端形成了粘著的基盤(basaldisk),用以附著在水草或其他物體上。群體生活的種類例如藪枝螅(Obelia)、反口端形成了匍匐莖(stolon)。水螅型的體壁是由兩層上皮肌肉細胞(epitheliomuscularcells)中間夾有一層很薄的中膠層(mesoglea)所組成,包圍中央的一個胃循環腔(gastrovascularcavity)。水螅類的胃循環腔為一簡單的空腔,也呈管狀,而珊瑚類的胃循環腔複雜,被體壁向心伸出的許多隔膜分隔成許多間隔,以增大消化及吸收的面積。 |
珊瑚綱Anthozoa
缽水母綱 Scyphozoa
十字水母綱 Staurozoa
立方水母綱 Cubozoa
Polypodiozoa
水螅綱 Hydrozoa
Trilobozoa (已滅絕)
刺胞動物除極少數種類為淡水生活外,絕大多數種均為海洋生活,多數在淺海,少數為深海種,現存約11000種。
刺胞動物不僅有細胞分化,而且開始分化出簡單的組織。刺胞動物的體壁由外胚層、內胚層和兩層細胞之間的中膠層構成。外胚層有4種細胞:上皮肌肉細胞、腺細胞、間細胞、刺細胞。
上皮肌肉細胞(epithelio-muscular cell)的特點是在上皮細胞內包含有肌原纖維。這種細胞具有上皮和肌肉的功能,所以稱為上皮肌肉細胞(epithelio-muscular cell)簡稱皮肌細胞。同時刺胞動物的上皮還具有像神經一樣的傳導功能,這是近些年來應用電生理學技術和電子顯微鏡檢來研究腔腸動物神經的一個發現。非神經的傳導(non-nervous conduction)或類神經(neuroid)傳導,首先是在腔腸動物得到證實的。上皮肌肉細胞既屬於上皮,也屬於肌肉的範圍。這表明上皮與肌肉沒有分開,是一種原始的現象。一般在上皮肌肉細胞的基部延伸出一個或幾個細長的突起,其中有肌原纖維(myofibrils),也有的上皮成分不發達,成為肌細胞(myocyte),有的是上皮成分發達,細胞呈扁平狀,肌原纖維呈單向排列,或者是2排肌原纖維呈垂直排列,也有的上皮成分發達呈圓柱狀,周圍有一系列的平滑肌環。肌纖維也分為橫紋肌、斜紋肌和平滑肌。每個肌原纖維都是由一束細絲組成,這些絲又分粗、細2種,與高等動物粗(肌球蛋白)、細(肌動蛋白)絲相似,其收縮機理也和高等動物的相似。關於肌肉的神經支配了解的不多,有的實驗證明,刺胞動物的神經與肌肉的接觸部分——神經肌肉突觸(neuromuscular synapses)的超微結構和神經肌肉連接(neuromus-cular junction),也都與高等動物的相似。
腺細胞(glandcell)是一種具分泌能力的上皮細胞。在水螅的基盤處及觸手的表皮層中腺細胞特別發達,它的分泌物可以幫助水螅體附著及捕食等。在具有骨骼的種類,例如水螅型群體及珊瑚類,其腺細胞可以分泌大量的角質或鈣質以形成幾丁質的圍鞘或鈣質外骨骼。胃層中也含有大量的腺細胞,細胞內含有大量的分泌顆粒,它可以轉化成消化酶,進行食物的細胞外消化。
間細胞(interstitialcell)位於上皮細胞之間,靠近中膠層處,是一些小型、圓形的細胞,單獨或成堆分佈,具大的細胞核,它是體內一種未分化的細胞,由它可以轉化成生殖細胞、刺細胞、腺細胞等其他類型的細胞。
刺細胞(cnidoblast)是刺胞動物特有的一種攻擊及防衛性細胞。在水螅類分佈於表皮層中,特別是在口區、觸手等部位,在缽水母及珊瑚類除了分佈於體表及觸手外,消化腔的胃絲、隔膜絲上也有大量的分佈以幫助捕食。刺細胞是一種特化了的上皮肌肉細胞,核位於基部,細胞頂端具一個刺針(cnidocil),伸出體表,其超微結構相似於鞭毛;刺的基部也有基粒。刺細胞內有一刺絲囊(nematocyst),囊的頂端為一蓋板(lid),囊內為細長盤卷的刺絲。當刺針或刺細胞受到刺激時,刺絲囊由刺細胞中被排出,同時刺絲也由刺絲囊外翻出來,形成不同長度的刺絲,用以捕食及防衛。實驗證明刺絲囊的排放機制是由機械刺激及化學刺激的聯合作用所引起,單獨地使用其中任何一種刺激並不引起排放。外界刺激作用於刺細胞,引起刺絲囊由周圍細胞質中吸收水分,改變囊壁滲透性,刺細胞隨之收縮,增加了刺絲囊內的壓力,刺絲衝破蓋板外翻出來而引起了排放。所以其排放是由於外界刺激直接作用於刺絲囊,而不是通過神經細胞。神經的傳導作用可能僅在大量的刺細胞的排放中起調節作用。已排放的刺絲囊其尖端不斷地滲出液體,這種液體對被捕物具有麻醉及毒殺作用。有人用一種海葵Aiptasia,從其消化道內胃絲上收集大量的刺絲囊毒液進行分析,結果發現其中有四種蛋白質成分。再用這四種蛋白質成分對招潮蟹(Uca)及喇咕蝦(Procambarus)做毒性實驗,結果發現其中有的蛋白質成分有毒殺作用,有的蛋白質有破壞細胞膜及神經索傳遞動作電位的能力,有的蛋白質成分引起實驗動物的強烈痙攣,這說明是通過神經系統的作用所引起。實驗結果說明由胃絲上收集的這些蛋白質具有神經毒素、肌肉毒素、溶血性及壞死性的特徵。有的刺胞動物其刺絲囊的毒素甚至對人也造成麻痹作用,例如大海蜇、霞水母等。每個刺細胞僅能排放一次,但可以由間細胞不斷地補充及更新,根據其排放出的刺絲囊及刺絲的形態、腔腸動物的刺絲囊有30多種,但每種動物一般有l~7種不等。例如水螅有四種,一種是穿刺刺絲囊(penetrant),用以穿刺並釋放毒液;一種是纏繞刺絲囊(volvent),這種不釋放毒液、但能纏繞捕獲物;還有兩種是粘著刺絲囊(glutinant),它們所排出的刺絲具有粘著及捕食功能。前兩種刺絲囊對化學刺激,特別是食物刺激比較敏感,后兩種對機械刺激敏感。有人觀察水螅在一次捕食時可排放出觸手上25%的刺絲囊,並在48小時內更新。
基本信息
- 中文名
- 刺胞動物門
- 別名
- 刺細胞動物門,腔腸動物門
- 拼音
- cì bāo dòng wù mén
- 拉丁學名
- Cnidaria
- 界
- 動物界
- 門
- 動物門
- 現狀
- 現多已廢棄不用
- 適用物種
- 刺胞動物及櫛水母動物