有孔蟲
形成於寒武紀時期的單細胞動物
有孔蟲是一類古老的原生動物,5億多年前就產生在海洋中,至今種類繁多。由於有孔蟲能夠分泌鈣質或硅質,形成外殼,而且殼上有一個大孔或多個細孔,以便伸出偽足,因此得名有孔蟲。有孔蟲是海洋食物鏈的一個環節,它的主要食物為硅藻以及菌類、甲殼類幼蟲等,個別種的食物是砂粒。有孔蟲是浮游生物中重要的組成部分,也是大多數海洋生物的重要的食物來源。
有孔蟲 Foraminifera
有孔蟲屬原生動物門肉足蟲綱有孔蟲亞綱,寒武紀到現在一直都存在著。是微小的真核單細胞動物。蟲體隱藏在細小的殼內,小殼分為許多小的內格,由一些微孔或孔狀接縫所穿通。殼一般是碳酸鈣質的,也有硅質和幾丁質的。某些有孔蟲為膠結殼。有孔蟲特別小,在鑽探過程中不會被破壞,對尋找石油特別有價值。蟲體由一團原生質構成。某些有孔蟲的殼(例如抱球蟲屬)大量堆積在現代洋底的許多地方,形成厚層沉積物。這種軟的、細粒的、被稱為抱球蟲軟泥。在過去地質時代也有與此相似的沉積,但已變成厚層的白堊和石灰岩。
有孔蟲
蟲體隱藏在細小的殼內,小殼分為許多小的內格,由一些微孔或孔狀接縫所穿通。殼一般是碳酸鈣質的,也有硅質的和幾丁質的。某些有孔蟲則是通過把細粒礦物質或其它外來的顆粒膠結起來,來建造自己的殼,這種殼為膠結的或含沙質的。由於有孔蟲特別小,在鑽探過程中不會被破壞,所以對尋找石油特別有價值。
某些有孔蟲可以很容易地用肉眼看見,少數種類的有孔蟲的殼,直徑達5厘米。某些有孔蟲的殼(例如抱球蟲屬)大量堆積在現代洋底的許多地方,形成厚層沉積物。這種軟的、細粒的、被稱為抱球蟲軟泥的鈣質物質,今覆蓋著約1.3億平方公里的海底。在地質時代也有與此相似的沉積,但已變成厚層的白堊和石灰岩。
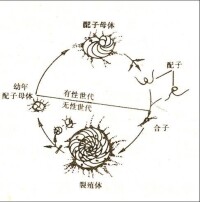
原生動物門(Protozoa)肉足蟲綱(Sarcodina)中的一個目。微小的真核單細胞動物。其大小一般在1mm以下,最大可達110mm。蟲體由一團原生質構成。原生質分化為兩層:外質和內質。具有一個由外質分泌物形成的或由分泌物膠結其它外來顆粒構築而成的殼(test)。殼是有孔蟲保存為化石的主要部分。生活環境以海洋為主。絕大多數都生活於正常海水環境中。僅有一個超科——瓶形蟲超科(Lagynacea)的某些代表生活於淡水環境,其薄而脆的殼,不易保存為化石。在海洋中部分生活於海底,部分漂浮於洋麵。生活史中存在著有性生殖和無性生殖交替進行的現象,稱為世代交替。同一個種中由於世代交替產生顯球型和微球型兩種不同類型的殼,叫做有孔蟲殼的雙型現象(dimorphism)。寒武紀到現代都有分佈。
晚古生代以來有孔蟲大體經歷了三個重大的繁盛期。
第一個繁盛期在石炭、二疊紀,蜓和內卷蟲類佔優勢。
第二個繁盛期在侏羅一白堊紀。浮遊類有孔蟲在中侏羅世開始出現,並在晚白堊世達到鼎盛。某些大型底棲有孔蟲類群的出現。
第三個繁盛期自始新世開始,是有孔蟲發展史上的全盛時代。
根足類有孔蟲目(Foraminiferida)的原生動物,特點是︰從藏在介殼內的單核或多核的細胞質體伸出細長的偽足。因種類不同,介殼大小可從極微小,到最大直徑超過5公分(2英寸)。且形狀和室的數目、化學組成以及表面定向亦因種而異。南太平洋種類的介殼特大,大洋洲島嶼的居民取作珠寶使用。埃及金字塔的始新世石灰岩中找到的錢幣蟲屬(Nummulite)介殼直徑大於5公分。有孔蟲類棲居於幾乎所有有保護物和適宜的食物(微小生物)的海域,幾乎在所有深度處都有發現。是現存浮遊和底棲的小型動物區系的重要組成部分。化石到處可見,因此可作為判定地質年代和探測石油的標準化石。有孔蟲死後,其石灰質空殼下沉,形成有孔蟲軟泥,覆蓋著約有30%的洋底面。石灰石和白堊是有孔蟲的海底沉積產物。
在英國南安普頓的海洋學中心研究海洋生物的Alan Hughes說,有孔蟲或許是海洋中除了細菌之外最豐富的海洋生物。大多數有孔蟲擁有碳酸鈣製成的複雜的殼。
但是在挑戰者深淵找到的有孔蟲缺少外殼。實際上,科學家撈到的85%的有孔蟲屬於軟殼的allogromiids——它們只佔海洋表面有孔蟲群的5%~20%。
這可能是因為海洋的最深處只有很少的碳酸鈣,這就意味著這些微生物可能無法製造外殼。“當你越走越深的時候,你就會到達一個叫做鈣補償點的深度,”Hughes解釋說。“在那個深度之下,你就開始發現軟殼的有孔蟲”。
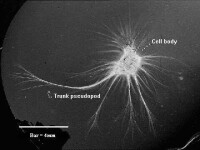
有孔蟲
有孔蟲
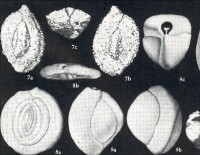
有孔蟲
包括以下構造:
一、殼室(chamber)。多個房室組成的殼其最早形成的、最小的房室叫做初房(proloculum),最後的一個叫終室。
二、口孔(aperture)。
三、隔壁(septum)。隔開相鄰兩個房室的殼壁叫隔壁。
四、縫合線(suture line)。隔壁與殼壁相交的線稱縫合線。
五、口面(oral face)。口孔周圍的殼壁稱為前壁或口面。
按有孔蟲的房室數目及排列方式殼有單房室殼(unilocular test)、雙房室殼(biloculartest)和多房室殼(multilocular test)
1. 單房室殼
由一個房室組成,房室上具一個或多個口孔。
2.雙房室殼
一般由一個球形的初房和一個管狀的第二房室組成,口孔常位於第二房室的末端。
3.多房室殼
由兩個以上的房室構成。可以分為以下幾種:
(1)單列式(uniserial)殼 房室的生長沿直線或弧形單列排列。
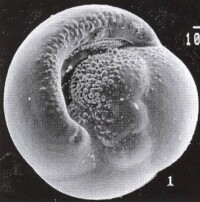
有孔蟲
有孔蟲種類繁多,包括1000多個屬、34000多種,現今種類約6000餘種,並且還以每天2個新種的速度飛快地增加,中國海約有1500餘種。有孔蟲數量豐富,分佈廣泛,可生活於各種各樣的海洋環境,其殼體可反映出非常有用的環境信息,作為環境指示生物可用於許多研究領域,被譽為“大海里的小巨人”。
有孔蟲亞種分類
從這些樣品中,北里洋的研究組發現了許多細菌和432種活的有孔蟲。後者的尺寸大約是數十微米。數百個有孔蟲擠在針尖大小的地方。絕大多數有孔蟲是軟殼的,它們或者是圓形,或者是針形,顏色是斑斕的棕色。他們在《科學》雜誌上報告了這個發現。
有孔蟲亞目分類
有孔蟲雕塑園
有孔蟲目根據最近的分類方案,下分6個亞目,19個超科
網足蟲(Allogromiina)
單房室假幾丁質殼。切片中呈薄膜狀,不分層,很少形成化石,常見於現代淡水和半鹹水沉積中。僅一個超科——瓶形蟲超科(Lagynacea),晚寒武世到現。
串珠蟲(Textulariina)
膠結殼,不分層,顆粒狀或纖維狀,有的具迷宮狀構造,據房室的多少分為兩個超科:
(1)砂盤蟲超科(Ammodiscacea)單房室或雙房室,常為球形、管狀、分枝狀或平旋、繞旋式殼。口孔簡單,無隔壁或具雛形隔壁。寒武紀至現代。
(2)曲杖蟲超科(Lituolacea)多房室,常為單列、雙列、三列式或螺旋式殼。隔壁發育完整。口孔單一或複雜。石炭紀至現代。
內卷蟲(Endothyrina)
鈣質微粒狀殼,殼壁二層或不分層。某些類別具假旋脊和小隔壁,殼壁內有時夾有少量外來物質,分兩個超科:
(1)擬砂戶蟲超科(Parathuramminacea)單房室,殼呈球形、管狀或串珠狀,殼壁常含外來物質,不分層,口孔簡單,有時可見雛形隔壁,奧陶紀至石炭紀;
(2)內卷蟲超科(Endothyracea)多房室,常為單列、雙列、平旋式殼,殼壁大多分為兩層,常具末端或基部口孔。志留紀至三疊紀。
蜓(Fusulinina)
三鄉“有孔蟲雕塑公園”
鈣質微粒狀殼,殼壁分層明顯,最多可達四層,旋脊、擬旋脊發育,隔壁常褶皺。石炭紀至二疊紀。僅一個超科—蜓超科(Fusulinacea)。
小粟蟲(Miliolina)
鈣質無孔似瓷質殼,常具假幾丁質內層,有時可含外來物質,以平旋式、繞旋式殼最為多見,口孔位於終室末端,常發育口孔飾變。石炭紀至現代。
僅一個超科-小粟蟲超科(Miliolacea)。
輪蟲(Rotaliina)
鈣質透明殼,殼壁有微孔,房室排列方式多樣,口孔類型複雜。中石炭世至現代。
共有12個超科。其中的抱球蟲超科(Globigerinacea)為浮遊有孔蟲,其它有孔蟲則營不同形式的底棲生活,通稱為底棲有孔蟲
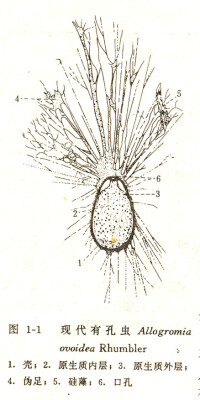
有孔蟲

有孔蟲

有孔蟲
現代的有孔蟲絕大多數都是海生的,只有少數生活在潟湖、河口等半鹹水的環境里,也有極少數廣鹽性的可以生活在超過正常鹽度的鹹水裡,還有極個別的可以生活在淡水裡,如瓶形蟲超科中的個別種屬。大多數的有孔蟲為底棲生活,少數為浮游生物。底棲的有孔蟲通常都可以在海底緩慢移動,只有少數固著生長。
影響有孔蟲生長、生殖和分佈的主要因素是水溫、深度和鹽分;適當食物的是否易得;基質的性質以及氧氣的供給等。現存的海洋有孔蟲種群由6個公認的不同動物區系組成;其中4個出現在較暖的海水中;兩個在較冷的海水中。
有孔蟲抵壓性
有孔蟲的生殖方式分有性生殖和無性生殖兩種。它們的生存空間與其它原生動物一樣甚為廣泛,分浮遊和底棲兩個類群。有孔蟲對環境的反應特別敏感,有明顯的深度分佈範圍,因而它們是最好的海深指示生物。由於不同時期有不同的有孔蟲,因此,根據有孔蟲的沉積物不但能確定地層的地質年代,而且還能提示地下情況,從而為尋找礦藏尤其是石油,提供重要依據。有孔蟲的個體極其微小,只有0.15毫米大小,需要藉助放大鏡才能分辨出來。它們比珊瑚在地球上出現的時間還早,而且一直繁衍至今。它們種類多繁殖力強,用無性生殖的生殖方式,一邊大量繁殖,一邊死亡,死後遺留下來的幾丁質、硅質和鈣質的殼沉積於礁石上與造礁珊瑚的骨骼膠結在一起。有的生物礁就是有孔蟲的遺骸佔據了主導地位。斐濟群島、埃利斯群島中的富納富提環礁,都是由有孔蟲的遺骸為主體構成的。
雖然有些種類只行無性生殖,而大部分種類具有規律的或偶見的有性世代。生殖過程通常需時1~3天。較小的種可在一個月完成有性和無性世代。但較大的種,往往需要1~2年。因為通常所有的細胞質均分配給新形成的幼體,因此生殖通常使母體的生命終止。
它們的家園遠在海平面之下,比珠穆朗瑪峰的高度還深。在太平洋最深的海溝——這個地球上最偏僻的角落發現了微小的生物。它們生活在馬里亞納海溝的挑戰者深淵底部的泥漿中。挑戰者深淵底部在西太平洋海面的11千米之下,那個深度的壓力是海面的1090倍。
在橫須賀的日本海洋科學技術機構的北里洋(Hiroshi Kitazato)領導了這項研究,他們使用一艘叫做海溝號(Kaiko)的潛艇收集了這個海溝的沉積物。這個遙控深海潛艇第一次潛入瑪里亞納海溝是在1995年。在2003年的一次任務中,它在海中失蹤。
死去的浮遊有孔蟲門生物不斷大量地向海床落下,它們蘊含豐富礦物的外殼以化石的形態在沉積物中被保留著。在1960年代起,一方面在深海鑽探計劃(Deep Sea Drilling Program)、海洋鑽探計劃(Ocean Drilling Program)及綜合海洋鑽探計劃(Integrated Ocean Drilling Program)的贊助,另一方面亦受到石油探采(oil exploration)的目的所推動下,由先進的深海鑽探技術所帶出的沉積物核心中,可以發現幾百萬年前的有孔蟲門生物化石。
大量的有品質素材
在其實際上無限量供應的化石外殼及其核心中有相對高準確度的時期抑制原型下,有孔蟲門生物能夠製造出非常高質素的浮遊有孔蟲門生物化石記錄,最早可以追溯至侏羅紀中期,其化石亦可以提供科學家一個不平衡的記錄去測試及記錄進化過程。因為化石記錄的極度高質素,令物種間的相互關係可以以此為基礎,能夠造成極詳細的影像。在很多情況下其後會在現存的樣本上以分子基因研究的方法作獨立確認。
在所有海洋環境中,有孔蟲是無組織無器官的單細胞生物。它們的個頭很小,只有幾毫米到幾十微米大。有孔蟲可以是浮游生物,也可以是底棲生物。浮遊有孔蟲生活在海洋的上部,然而底棲有孔蟲一般生活在洋底或洋底下面。
有孔蟲經常被用於生物地層研究或古環境重建。底棲有孔蟲能很好的代表海洋的深度,所以在古海洋研究中經常使用有孔蟲進行放射性碳測年。在開闊海洋中發現的浮遊有孔蟲經常被用于海洋洋流或氣候研究。研究人員也會利用有孔蟲從事海洋污染研究。
有孔蟲群落包含豐富的環境信息,對水體環境變化響應敏銳,而底棲有孔蟲對反映大洋底層水團具有明顯優勢。中國科學院南京地質古生物研究所李保華研究員團隊的郭啟梅博士與韓國、葡萄牙等國的科學家合作,對綜合大洋鑽探(IODP)339航次U1391站位1.3Ma以來的沉積物進行了高解析度分析。通過底棲有孔蟲系統古生物學與定量統計研究,結合精確同位素地層年代建立,依據底棲有孔蟲特徵組合("Elevated Epibenthos" group)、關鍵分子Planulina ariminensis和同位素組成變化,揭示了長時間尺度上地中海溢出流的詳細演化過程。研究成果發表於2021年1月《第四紀科學評論》(Quaternary Science Reviews)上。