基因轉錄
基因轉錄
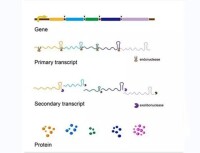
基因轉錄是在細胞核和細胞質內進行的。它是指以DNA的一條鏈為模板,按照鹼基互補配對原則,合成RNA的過程。基因轉錄有正調控和負調控之分。
如細菌基因的負調控機制是當一種阻遏蛋白(repressor protein)結合在受調控的基因上時,基因不表達;而從靶基因上去除阻遏蛋白后,RNA聚合酶識別受調控基因的啟動子,使基因得以表達,這是正調控。這種阻遏蛋白是反式作用因子。
基因轉錄
這篇文章的試驗方法是,通過高密度的寡核苷酸晶元,反映出人21和22號染色體的幾乎所有的非重複序列,通過這種晶元,檢測三種轉錄因子,Sp1、 cMyc、和p53的結合位點。結果表明,每種轉錄因子都有大量的TFBS與之結合。然而,只有22%的轉錄因子結合位點分佈在蛋白編碼基因的5'端, 36%的TFBS分佈在蛋白編碼基因的中部或3'端,並且這36%的TFBS常常和基因組中的非蛋白編碼RNA分佈在一起。這暗示,在人的基因組中,不僅包含蛋白編碼基因,也包含數量相當的非編碼基因(noncoding genes),他們都受常見的轉錄因子所調控。
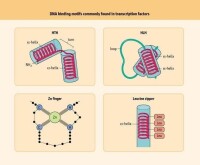
真核生物在轉錄時往往需要多種蛋白質因子的協助。一種蛋白質是不是轉錄機構的一部分往往是通過體外系統看它是否是轉錄起始所必須的。一般可將這些轉錄所需的蛋白質分為三大類:
(2)某些轉錄因子能與RNA聚合酶結合形成起始複合物,但不組成遊離聚合酶的成分。這些因子可能是所有啟動子起始轉錄所必須的。但亦可能僅是譬如說轉錄終止所必須的。但是,在這一類因子中,要嚴格區分開哪些是RNA聚合酶的亞基,哪些僅是輔助因子,是很困難的。
(3)某些轉錄因子僅與其靶啟動子中的特異順序結合。如果這些順序存在於啟動子中,則這些順序因子是一般轉錄機構的一部分。如果這些順序僅存在於某些種類的啟動子中,則識別這些順序的因子也只是在這些特異啟動子上起始轉錄必須的。
基因轉錄
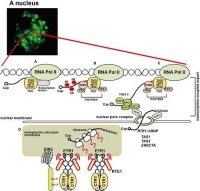
在真核生物中,DNA的轉錄在細胞核中進行,其中rRNA的合成發生在核仁,mRNA的tRNA的合成發生在核質中。
在原核生物中,轉錄在細胞質的核質區進行。
轉錄開始不需要引物,鏈的延長方向也是 5′→ 3′。
每次被轉錄的DNA只是一個小區段,而且是其中的一條鏈。
我們將用作RNA合成的模板的鏈叫做反義鏈;另一條不做模板的鏈叫有義鏈。
對於整個DNA雙鏈,每條鏈上有的區段用作有義鏈,有的區段用作反義鏈。
3. 原核生物參與轉錄的酶
RNA聚合酶
有五種亞基:a、b、b′、w、s,此外每個酶分子還含有2個Zn原子。
| a2bb'ws | = | a2bb'w + s |
| 全酶 | 核心酶 |
s亞基:用於識別DNA上轉錄的起始位點,引導核心酶結合到它上面
核心酶:由s亞基識別起始點后,由核心酶負責解開DNA雙鏈、RNA鏈的延伸、恢復後面的DNA雙螺旋
a亞基 —— 與啟動子結合
b亞基 —— 催化磷酸二酯鍵的形成
b'亞基 —— 與DNA模板結合
(1)轉錄的啟動
DNA上存在著轉錄的起始信號,它是特殊的核苷酸序列,稱為啟動子。
轉錄是由RNA聚合酶全酶結合於啟動子而被啟動的。
基因轉錄
(2)轉錄的起始
當聚合酶結合到啟動子上后,在啟動子附近將DNA局部解鏈,約解開17個鹼基對。(酶與啟動子結合的部位是AT富集區,有利於解鏈)
第一個核苷三磷酸(常常是GTP或ATP)結合到全酶上,形成“啟動子-全酶-核苷三磷酸”三元起始複合物。
第二個核苷酸參入,連結到第一個核苷酸的3'羥基上,形成了第一個磷酸二酯鍵。
s因子從全酶上掉下,又去結合其它的核心酶。
(3)鏈的延伸
當s因子從核心酶上脫落後,核心酶與DNA鏈的結合變得疏鬆(依靠其蛋白質的鹼性與酸性核酸之間的非特異性的靜電引力),可以在模板鏈上滑動,方向為DNA模板鏈的 3′→ 5′,同時將核苷酸逐個加到生長的RNA鏈的3'-OH端,使RNA鏈以 5′→ 3′方向延伸。
新合成的RNA鏈能與模板形成RNA-DNA雜交區,這個雜交區也在隨著RNA聚合酶的移動而不斷地移動著。
(4)轉錄的終止
DNA分子上有終止轉錄的特殊信號,也是特定的核苷酸序列,稱為終止子。
RNA聚合酶可以識別終止子,它在一種蛋白質 —— r因子的幫助下,終止轉錄,放出RNA鏈;有時,RNA聚合酶不需要r因子的幫助即可終止轉錄。
核心酶釋放了RNA后,也離開DNA。
DNA上的解鏈區重新形成雙螺旋。
基本信息
- 中文名
- 基因轉錄
- 外文名
- Gene transcription