頭索動物亞門
頭索動物亞門
頭索動物亞門(Cephalochordata)又稱無頭動物(Acrania),是脊索動物門的一個亞門。身體似魚但無真正的頭,終身都有一條縱貫全身的脊索,背側有神經管,咽部具許多條鰓裂。其典型代表種文昌魚(Branchostoma)。頭索動物亞門包括文昌魚等30餘種海棲魚形動物。種類雖不多,僅有頭索綱一綱,但在動物學上卻佔有重要地位,由於它們的身上以簡單的形式終生保留著脊索動物的三大基本特徵,長期以來為研究脊索動物起源的學者所重視。在動物學研究中佔有重要地位。

頭索動物亞門
沿背側有一縱行皮膚褶皺,稱為背鰭;再后延至尾部高起,成為尾鰭,作矛形。自尾鰭腹面向前至體后約1/3處的皮膚褶襞,稱為腹褶(metapleure)。左右兩腹褶向後的匯合處是為腹孔,或圍鰓腔孔(abdominal pore),腹鰭與尾鰭之間的小孔,為肛門。頭部前端突出部位為吻。其後的腹面有一環形薄膜,成為口笠。其邊緣生有36-50條觸鬚。口笠的內腔稱為前庭。
神經管的形成:原腸胚背中線的外胚層細胞向內凹陷,形成神經板,其兩同時翹起形成一對縱褶,叫神經褶。神經褶又相向生長,在神經板背側癒合;神經板 兩側亦向上包圍生長;彼此接合。於是在背面出現一條中空的管道,即神經管。
脊索與中胚層的發生:在神經管形成的同時,原腸腔的背面、神經板下方的內胚層細胞向背方突出,逐漸脫離內胚層形成一條縱貫的脊索;在脊形成的同時,脊索突 起的兩側也發生若干成對的分節狀突起,最後脫離內胚層,自成一腔囊,叫體腔囊,也以是中胚層。將來發育成為體腔。內胚層除了分離成脊索和中胚層外,其他部分則形成腸管的內壁。
因具有縱貫體軀一直達到前端的脊索而得名;頭無明顯分化,與體軀不能截然分開,故又名無頭類,通稱文昌魚、矛形魚。頭索動物終生具有縱貫背部,起支撐作用的脊索;有背神經管和咽鰓裂。這些在高等脊索動物中只存在於胚胎或幼蟲期,成體消失或分化為更高級的器官。

頭索動物亞門
這類動物通常少活動,大部時間將身體埋在泥沙中,僅以前端露出沙外,借水流帶食物進入口中。它們的脊索縱貫全身,且伸延到神經管的前面,故稱頭索動物,或全索動物。又因沒有真正的頭部,因此又稱無頭類(Acrania)。本亞門僅有一綱,即頭索綱。是一類終生具有發達脊索、背神經管和咽鰓裂等特徵的無頭魚形脊索動物。脊索不但終生保留,且延伸至背神經管的前方,故稱頭索動物。又因本亞門動物都缺乏真正的頭和腦,所以又稱無頭類。頭索動物分佈很廣,遍及熱帶和溫帶的淺海海域,其中尤以北緯48°至南緯40°之間的沿海地區數量較多。它們對底質要求比較嚴格,通常僅局限在有機質含量低的純凈粗砂和中砂中。生活時身體埋入砂中,僅前端外露,用以進行呼吸和濾食水體中的硅藻。綱內共有二屬:文昌魚屬(Branchiostoma)和偏文昌魚屬(Asymmetron)。文昌魚屬的體長約5厘米,但產於美國西海岸的加州文昌魚體長達10厘米,廣泛分佈在世界熱帶和亞熱帶的沿岸海域以至暖溫帶地區。偏文昌魚體形較高,體長一般為16毫米,明顯地小於文昌魚,形態特徵是只在身體右側具生殖腺,分佈僅限於印度洋—太平洋熱帶海區,中國北部灣西北部和汕頭附近的淺海區曾有發現。
起源原索動物沒有留下化石,但有一個例外。在加拿大不列顛哥倫比亞的著名的布爾吉斯頁岩中發現了寒武紀中期的一種動物的化石,命名為Pikaia gracilens(皮克魚),魚形,約5cm,具有脊索和“》”形分節肌節,與文昌魚很相似。毫無疑問是一種脊索動物,比最早的脊椎動物化石還早幾百萬年。但在缺乏其他的相關化石的情況下,不大可能決定它與最早的脊椎動物的關係。1999年11月《Nature》雜誌上發表了中國學者舒德干等人在5.3億年前澄江動物群化石中發現的“華夏鰻“的文章,華夏鰻十分像現代的文昌魚。皮克魚比澄江動物群晚1000萬年。地位⑴祖先可能是原始的無頭類,與無脊椎動物有共同祖先;⑵由於適應不同生活方式而演變為兩支,一支演變為原始有頭類,導向脊椎動物進化之路,另一支特化為旁支,演變成頭索動物的鰓口科動物;⑶認為頭索動物是當前脊椎動物的原始類群,是脊椎動物的姐妹群。在動物學上佔有重要的地位。
文昌魚類是本亞門的主要類群。
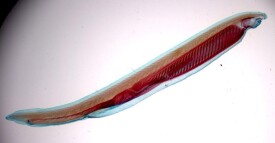
外形文昌魚的體形略似小魚,無明顯的頭部,左右側扁,半透明,可隱約見到皮下的肌節(myomere)和腹側塊狀的生殖腺;身體兩端尖出,故有雙尖魚(Amphioxus)之稱,又因其尾形很像矛頭而名海矛。一般體長約50mm,但產於美國的加州文昌魚(Branchiostomacaliforniense)可超過100mm,是該屬中已知個體最大的一種。前端的腹面為一漏斗狀的口笠(oral hood),口笠內為前庭(vestibule),內壁有輪器(wheel organ),由前庭引向位於一環形緣膜(velum)中央的口。口笠和緣膜的周圍分別環生觸鬚(cirri)及緣膜觸手(velar tentacle),具有保護和過濾作用,可阻擋粗砂等物隨水流進入口中。整個背面沿中線有一條低矮的背鰭(dorsalfin),往後與高而繞尾的尾鰭(caudal fin)相連。此外在肛門之前還有肛前鰭(preanal fin)。無偶鰭,只在身體前部的腹面兩側各有一條由皮膚下垂形成的縱褶,稱為腹褶(metapleura fold)。腹褶和肛前鰭的交界處有一腹孔(atripore),是咽鰓裂排水的總出口,故又名圍鰓腔孔。生活方式文昌魚喜棲淺海水質清澈的沙灘上,平時很少活動,常把身體半埋於沙中,前端露出沙外,或者左側貼卧沙面,借水流攜帶矽藻等浮游生物進入口內。夜間較為活躍,憑藉體側肌節的交錯收縮,在海水中作短暫的游泳。壽命約2年8個月左右。5~7月為生殖季節,一生中可繁殖3次,其中以最後一次產卵最多。福建沿海海區曾是中國文昌魚生息繁衍的理想場所。1932-1956年,文昌魚在同安縣一地的捕撈區面積約有22km2,每升沙樣中文昌魚的數量達357條,當時因年產量約57噸而高居世界首位。1956年,由於廈門高(崎)集(美)海堤建成后,海況(切斷了環島的海流、沙地面積大幅度減少、鹽度降低、水流減緩、淤泥沉積等)發生很大變化,再加上圍海造田和亂捕濫撈等原因,嚴重地破壞了文昌魚的生態環境,因而造成數量急劇下降的惡果,年產量減少了40%。1970年,當地的收購量甚至降低到只有1t左右。1986年,文昌魚在這一海域的分佈區已縮小到0.5km2,而每升沙樣中的數量進一步減少到3條,使原來名聞遐邇的文昌魚產地,在20世紀半個世紀內變得完全失去了水產捕撈價值。內部構造特徵1.皮膚:皮膚薄而半透明,由單層柱形細胞的表皮和凍膠狀結締組織的真皮兩部分構成,表皮外覆有一層角皮層(cuticle)。表皮外在幼體期生有纖毛,成長后則消失殆盡。2.骨骼:文昌魚尚未形成骨質的骨骼,主要是以縱貫全身的脊索作為支持動物體的中軸支架。脊索外圍有脊索鞘膜,並與背神經管的外膜、肌節之間的肌隔、皮下結締組織等連續。脊索細胞呈扁盤狀,其超顯微結構與雙殼類軟體動物的肌細胞比較相似,收縮時可增加脊索的硬度。此外,在口笠觸鬚、緣膜觸手、輪器內部也都有角質物支持,奇鰭和鰓裂的鰭條(fin rays)及鰓條(gill bar)由結締組織支持。3.肌肉:文昌魚背部的肌肉厚實而腹部比較單薄,與無脊椎動物周身體壁厚薄均勻的情況不同。全身主要的肌肉是60多對按節排列於體側的"V"字形肌節(myomere),尖端朝前,肌節間被結締組織的肌隔(myocomma)所分開。兩側的肌節互不對稱,便於文昌魚在水平方向作彎曲運動。此外,還有分佈在圍鰓腔腹面的橫肌和口緣膜上的括約肌等,控制圍鰓腔的排水及口孔的大小。4.消化和呼吸器官:文昌魚靠輪器和咽部纖毛的擺動,使帶有食物微粒的水流經口入咽,食物被濾下留在咽內,而水則通過咽壁的鰓裂至圍鰓腔,然後由腹孔排出體外。作為收集食物和呼吸場所的咽部極度擴大,幾乎佔據身體全長的1/2,咽腔內的構造與尾索動物相似,也具有內柱、咽上溝和圍咽溝等。文昌魚幼體的鰓裂直接開口於體表,後來形成圍鰓腔,以腹孔作為咽部鰓裂的總出水口。咽內的食物微粒被內柱細胞的分泌物粘結成團,再由纖毛運動使它從後向前流動,經圍咽溝轉到咽上溝,往後推送進入腸內。腸為一直管,向前伸出一個盲囊,突入咽的右側,稱為肝盲囊(hepatic diverticulum),能分泌消化液,與脊椎動物的肝臟為同源器官。食物團中的小微粒可進入肝盲囊,被肝盲囊細胞所吞噬,營細胞內消化,大微粒在腸內分解成小微粒后,也轉到肝盲囊中進行細胞內消化,未消化的物質由肝盲囊重返腸中,在後腸部進行消化和吸收。腸的末端開口於身體左側的肛門。咽腔是文昌魚完成呼吸作用的部位。咽壁兩側有60多對鰓裂,彼此以鰓條分開,鰓裂內壁布有纖毛上皮細胞和血管。水流進入口和咽時,藉纖毛上皮細胞的纖毛運動,通過鰓裂,並使之與血管內的血液進行氣體交換,最後,水再由圍鰓腔經腹孔排出體外。有人認為文昌魚纖薄的皮膚也具有直接從水中攝取氧氣的能力。5.血液循環:文昌魚循環系統屬於閉管式,即血液完全在血管內流動,這種情形與脊椎動物基本相同。無心臟,但是具搏動能力的腹大動脈(ventral aorta),因而被稱為狹心動物。由腹大動脈往兩側分出許多成對的鰓動脈(branchial arteries)進入鰓隔,鰓動脈不再分為毛細血管,它在完成氣體交換的呼吸作用后,於鰓裂背部匯入2條背大動脈根。背大動脈根內含多氧血,往前流向身體前端,向後則由左、右背大動脈根合成背大動脈(dorsal aorta),再由此分出血管到身體各部。血液無色,也沒有血細胞和呼吸色素,動脈中的血液通過組織間隙進入靜脈。從身體前端返回的血液通過體壁靜脈(parietal vein)注入一對前主靜脈(anterior cardinal vein);尾的腹面有一條尾靜脈(caudal vein),收集一部分身體後部回來的血液,進入腸下靜脈(subintestinal vein),大部分血液則流進2條後主靜脈(posterior cardinal vein)。左、右前主靜脈和兩條後主靜脈的血液全部匯流至一對橫形的總主靜脈(common cardinal vein),或稱居維葉氏管(ducts Cuvieri)。左、右總主靜脈會合處為靜脈竇(sinus venosus),然後通入腹大動脈。從腸壁返回的血液由毛細血管網集合成腸下靜脈,尾靜脈的部分血液也注入其中;腸下靜脈前行至肝盲囊處血管又形成毛細管網,由於這條靜脈的兩端在肝盲囊區都形成毛細血管,因此稱作肝門靜脈(hepatic portal vein)。由肝門靜脈的毛細血管再一次合成肝靜脈(hepatic vein)並將血液匯入靜脈竇內。
基本信息
- 中文名
- 頭索動物亞門
- 外文名
- Amphioxus
- 別名
- 無頭動物
- 拉丁學名
- Cephalochordata
- 界
- 動物界
- 亞門
- 頭索動物亞門
- 門
- 體征
- 身體似魚但無真正的頭,終身都有一條縱貫全身的脊索,背側有神經管,咽部具許多條鰓裂
- 代表種
- Branchostoma(文昌魚)